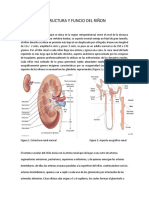
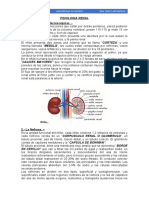
You might also like
- Atencion de Enfermeria Al Paciente Renal Corregido ANATOMIA Y FISIOLOGIADocument100 pagesAtencion de Enfermeria Al Paciente Renal Corregido ANATOMIA Y FISIOLOGIAJosé RamirezNo ratings yet
- Coagulación Para Todos: Medicina Para TodosFrom EverandCoagulación Para Todos: Medicina Para TodosRating: 4.5 out of 5 stars4.5/5 (3)
- Instituto Superior Tecnológico Cruz Roja EcuatorianaDocument7 pagesInstituto Superior Tecnológico Cruz Roja EcuatorianaAngelicaEmoxaNo ratings yet
- Anatomia y Fisiología del Sistema Nervioso IIFrom EverandAnatomia y Fisiología del Sistema Nervioso IIRating: 4 out of 5 stars4/5 (28)
- Material de Apoyo Al Estudio Del Aparato UrinarioDocument19 pagesMaterial de Apoyo Al Estudio Del Aparato UrinarioEsmeraldaNo ratings yet
- Anatomía y Fisiología del Sistema NerviosoFrom EverandAnatomía y Fisiología del Sistema NerviosoRating: 4 out of 5 stars4/5 (19)
- Aparato Urinario Marco TeòricoDocument18 pagesAparato Urinario Marco TeòricoAlxAgorNo ratings yet
- AparatoUrinario12Document15 pagesAparatoUrinario12Karina Ambriz ParedesNo ratings yet
- Aparato UrinarioDocument11 pagesAparato UrinarioDariiana Benavides100% (1)
- Resumen UrinarioDocument5 pagesResumen UrinarioValeria Belén SantanderNo ratings yet
- Apuntes Nefro (Principal)Document120 pagesApuntes Nefro (Principal)Crixalis 145No ratings yet
- Estructura de Los RiñonesDocument16 pagesEstructura de Los RiñonesKathy AmoradiosNo ratings yet
- Anatomía Del RiñónDocument4 pagesAnatomía Del RiñónMaite PashmayNo ratings yet
- Sistema UrinarioDocument8 pagesSistema UrinarioAlexis S. RuizNo ratings yet
- Nefrectomia, Trabajo FinalDocument15 pagesNefrectomia, Trabajo FinalJose Lopez De [OssioNo ratings yet
- Anatomia y Fisiologia Renal para EnfermerasDocument172 pagesAnatomia y Fisiologia Renal para EnfermerasMaria Camila Ramírez Gutiérrez100% (1)
- Tema 4 AP. Urinario AnatomíaDocument11 pagesTema 4 AP. Urinario AnatomíaJuanjo MedinaNo ratings yet
- RIÑÓNDocument7 pagesRIÑÓNParker GreenNo ratings yet
- RiñónDocument14 pagesRiñónJose Alberto Lazcano BrunoNo ratings yet
- Sistema Urinario Histologia Uabp 1Document7 pagesSistema Urinario Histologia Uabp 1lua robertiNo ratings yet
- Histología Del RiñonDocument6 pagesHistología Del RiñonKarime LopezavilaNo ratings yet
- NefrologíaDocument59 pagesNefrologíakarlasifontesNo ratings yet
- Tejido MuscularDocument9 pagesTejido MuscularDaniela HeLoNo ratings yet
- Cs. Biológicas - Sist. RenalDocument59 pagesCs. Biológicas - Sist. RenalAna belen GomezNo ratings yet
- NEFRONADocument14 pagesNEFRONANadia Sabine Soler AlarcónNo ratings yet
- Copia de Portada Tesis Tercer SemestreDocument19 pagesCopia de Portada Tesis Tercer SemestreMariana RoblesNo ratings yet
- Fisiología RenalDocument20 pagesFisiología RenalAngel GUNo ratings yet
- RIÑONESDocument9 pagesRIÑONESEstibaliz Carmona BerriosNo ratings yet
- Morfofisiologia Humana IIDocument18 pagesMorfofisiologia Humana IIDanna TorrellesNo ratings yet
- Aparato UrinarioDocument64 pagesAparato UrinarioSalvador BARRON MELONo ratings yet
- Histologia - Resumen de Aparato UrinarioDocument11 pagesHistologia - Resumen de Aparato UrinarioValeria MéndezNo ratings yet
- Fisiologia y Fisiopatologia RenalDocument20 pagesFisiologia y Fisiopatologia RenalAnnibel Flores ReyesNo ratings yet
- Estructura y Funcio Del RiñonDocument4 pagesEstructura y Funcio Del RiñonKEILA INTERNETNo ratings yet
- La Nefrona, Tubulos Proximales y DistalesDocument2 pagesLa Nefrona, Tubulos Proximales y DistalesHeidi Esther BaezNo ratings yet
- Tarea 2 Primer Parcial UroanalisisDocument8 pagesTarea 2 Primer Parcial UroanalisisAmdres AlmonteNo ratings yet
- Apuntes Aparato UrinarioDocument12 pagesApuntes Aparato UrinarioPaz GarciaNo ratings yet
- Fisiologia RenalDocument20 pagesFisiologia RenalMonique GloverNo ratings yet
- Anatomía y fisiología del sistema renalDocument15 pagesAnatomía y fisiología del sistema renalgomezNo ratings yet
- FISIOLOGIA RENALDocument21 pagesFISIOLOGIA RENALDayana Ruiz HerreraNo ratings yet
- Aparato Urinario 199910 DownloableDocument8 pagesAparato Urinario 199910 DownloableFrank FQNo ratings yet
- AparatoUrinario7págDocument8 pagesAparatoUrinario7págFrank FQNo ratings yet
- Capitulo Ii RiñonDocument36 pagesCapitulo Ii Riñonadrianamargarita0007.amooNo ratings yet
- AnatomíaDocument66 pagesAnatomíaMarìa Daniela ZambranoNo ratings yet
- Tema 10. Aparato UrinarioDocument14 pagesTema 10. Aparato UrinarioanahitamarNo ratings yet
- Los RiñonesDocument8 pagesLos RiñonesPedro Jose Felipa BazalarNo ratings yet
- Anatomía y Fisiología Del Riñón, Formación de Orina, Equilibrio Hidroelectrolític0 y Ácido BaseDocument19 pagesAnatomía y Fisiología Del Riñón, Formación de Orina, Equilibrio Hidroelectrolític0 y Ácido BasejllontopllaNo ratings yet
- Anatomia de Los RiñonesDocument6 pagesAnatomia de Los RiñonesAlexandre QuisbertNo ratings yet
- Elementos de La Función RenalDocument8 pagesElementos de La Función RenalFlorencia BarsiNo ratings yet
- Generalidades de Los RiñonesDocument12 pagesGeneralidades de Los RiñonesMariaAlexandraPinoNo ratings yet
- Manual de exploración urinariaDocument80 pagesManual de exploración urinariacampsar50% (2)
- ANATOMIADocument6 pagesANATOMIAMontserrat ZaldoNo ratings yet
- Principal UroDocument253 pagesPrincipal UroAriel GuerreroNo ratings yet
- histologia riñones puntosDocument13 pageshistologia riñones puntosgabriela escobarNo ratings yet
- Unidad 6. El RiñonDocument10 pagesUnidad 6. El RiñonFederico Pennacchioni polaNo ratings yet
- Clase de RenalDocument34 pagesClase de RenalDeniseDelPuertoNo ratings yet
- Apuntes BiologiaDocument4 pagesApuntes BiologiaMartin Paredes SanchezNo ratings yet
- Anatomia y Fisiologia Renal Del Riñon - ResumenDocument8 pagesAnatomia y Fisiologia Renal Del Riñon - ResumenEstephany MolinaNo ratings yet
- Patologías Del Aparato Genito-Urinario PDFDocument94 pagesPatologías Del Aparato Genito-Urinario PDFEster SadornilNo ratings yet
- Fisiologia RenalDocument13 pagesFisiologia RenalDelmira Andia challcoNo ratings yet
- Programación 2010A Fisiología Grupo 02Document7 pagesProgramación 2010A Fisiología Grupo 02ehtoledoc2127100% (1)
- Generalidades Sistema NerviosoDocument5 pagesGeneralidades Sistema Nerviosoehtoledoc2127100% (2)
- Programación 2010A Farmacología Grupo 02Document5 pagesProgramación 2010A Farmacología Grupo 02ehtoledoc2127No ratings yet
- Programación 2010A Fisiología Grupo 03Document7 pagesProgramación 2010A Fisiología Grupo 03ehtoledoc2127No ratings yet
- Sistema Respiratorio en RedDocument65 pagesSistema Respiratorio en RedehtoledocNo ratings yet
- Programación 2010A Fisiología Grupo 03Document7 pagesProgramación 2010A Fisiología Grupo 03ehtoledoc2127No ratings yet
- Programación 2010A Farmacología Grupo 03Document4 pagesProgramación 2010A Farmacología Grupo 03ehtoledoc2127No ratings yet
- Programación 2010A Fisiología Grupo 01Document7 pagesProgramación 2010A Fisiología Grupo 01ehtoledoc2127No ratings yet
- Programación 2010A Fisiología Grupo 01Document7 pagesProgramación 2010A Fisiología Grupo 01ehtoledoc2127No ratings yet
- Programación 2010A Fisiología Grupo 02Document7 pagesProgramación 2010A Fisiología Grupo 02ehtoledoc2127No ratings yet
- Programación 2010A Farmacología Grupo 01Document5 pagesProgramación 2010A Farmacología Grupo 01ehtoledoc21270% (1)
- Red Corazon MiaDocument64 pagesRed Corazon MiaehtoledocNo ratings yet
- Programación 2010A Farmacología Grupo 02Document5 pagesProgramación 2010A Farmacología Grupo 02ehtoledoc2127No ratings yet
- Programación 2010A Farmacología Grupo 01Document5 pagesProgramación 2010A Farmacología Grupo 01ehtoledoc21270% (1)
- Programación 2010A Fisiología Grupo 01Document7 pagesProgramación 2010A Fisiología Grupo 01ehtoledoc2127No ratings yet
- Programación 2010A Fisiología Grupo 01Document7 pagesProgramación 2010A Fisiología Grupo 01ehtoledoc2127No ratings yet
- Sistema Circulatorio Corazón y MiaDocument12 pagesSistema Circulatorio Corazón y Miaehtoledoc100% (8)
- Fisiología Del Aparato RespiratorioDocument15 pagesFisiología Del Aparato Respiratorioehtoledoc212767% (3)
- Causa EfectoDocument8 pagesCausa Efectoehtoledoc2127No ratings yet
- Programación 2009 FarmacologíaDocument14 pagesProgramación 2009 Farmacologíaehtoledoc2127100% (1)
- Sistema Renal UCC RedDocument42 pagesSistema Renal UCC Redehtoledoc100% (2)
- Causa Efecto FarmacologíaDocument8 pagesCausa Efecto Farmacologíaehtoledoc2127100% (1)
- Sistema Circulatorio Corazón y MiaDocument12 pagesSistema Circulatorio Corazón y Miaehtoledoc100% (8)
- Sistema Circulatorio SangreDocument4 pagesSistema Circulatorio Sangreehtoledoc2127100% (1)
- Programación 2009 FisiologíaDocument25 pagesProgramación 2009 Fisiologíaehtoledoc2127100% (1)
- 2009 Grupos y Acciones FarmacológicosDocument12 pages2009 Grupos y Acciones Farmacológicosehtoledoc212783% (6)
- 2008 FarmacodinamiaDocument4 pages2008 Farmacodinamiaehtoledoc2127100% (1)
- Fisiología Muscular BlogDocument79 pagesFisiología Muscular Blogehtoledoc2127100% (3)
- 2008 FarmacocinéticaDocument9 pages2008 Farmacocinéticaehtoledoc2127100% (5)
- Puerta ContraplacadaDocument16 pagesPuerta ContraplacadaBrayan Farfan ValladolidNo ratings yet
- Practica 10Document48 pagesPractica 10Erick Vega QuispeNo ratings yet
- Leche en polvo: proceso de secado por atomizaciónDocument44 pagesLeche en polvo: proceso de secado por atomizaciónAJ CarlosNo ratings yet
- Diseño Redes Acceso Fijo PDFDocument18 pagesDiseño Redes Acceso Fijo PDFJhonny Santos Moran0% (1)
- Diseno Neumatico y Electronico Del Sistema de ControlDocument0 pagesDiseno Neumatico y Electronico Del Sistema de ControlVíctor VargasNo ratings yet
- Smcadena - 1. Unidades de MedidaDocument42 pagesSmcadena - 1. Unidades de Medidaandres pedrazaNo ratings yet
- Tema 3 Análisis PVTDocument36 pagesTema 3 Análisis PVTpabloNo ratings yet
- Exposicion Hornos de FundicionDocument42 pagesExposicion Hornos de FundicionRoberto Fernandez AsencioNo ratings yet
- FEUM 12a Ed. Sistemas Criticos Recuperad PDFDocument100 pagesFEUM 12a Ed. Sistemas Criticos Recuperad PDFsnrubicsNo ratings yet
- Cido Acetil SalicilicoDocument6 pagesCido Acetil SalicilicoDiego Morales MataNo ratings yet
- Prácticas de Física 1Document240 pagesPrácticas de Física 1alvamoch2100% (7)
- Tecno InformeDocument25 pagesTecno InformeM C Ch AragonNo ratings yet
- Construcciones PronominalesDocument4 pagesConstrucciones PronominalesUniformesForjadorNo ratings yet
- Informe Convalidación de Practica ProfesionalDocument17 pagesInforme Convalidación de Practica ProfesionalCristian Alejandro Osses Gonzalez0% (1)
- Factores de conversión, densidad y estructura atómicaDocument29 pagesFactores de conversión, densidad y estructura atómicaGiovanny IsazaNo ratings yet
- Trabajo GasesDocument3 pagesTrabajo GasesSheila100% (1)
- CXS 221sDocument5 pagesCXS 221sandres felipe mendoza obandoNo ratings yet
- Tipos de RocaDocument4 pagesTipos de Rocadroopydoopy3No ratings yet
- Historia y evolución de los biomaterialesDocument13 pagesHistoria y evolución de los biomaterialesAnnelle PalafoxNo ratings yet
- Trabajo Final Contaminacion Aire-Doctorado-2011 (Grupo1)Document117 pagesTrabajo Final Contaminacion Aire-Doctorado-2011 (Grupo1)Juan SalcedoNo ratings yet
- INFILTRACIONDocument20 pagesINFILTRACIONNicolas RomaniNo ratings yet
- Ensayo Sobre La Historia de La QuimicaDocument9 pagesEnsayo Sobre La Historia de La QuimicaAngie V. MatamorosNo ratings yet
- Desajuste de PotenciaDocument5 pagesDesajuste de PotenciaAnonymous UvNIhx5No ratings yet
- Clase BioenergéticaDocument31 pagesClase BioenergéticaIvan AguilarNo ratings yet
- MATRIZ-IDENTIFICACIÓN-RIESGOSDocument6 pagesMATRIZ-IDENTIFICACIÓN-RIESGOSNicolassalgueroNo ratings yet
- Enlace IónicoDocument6 pagesEnlace IónicoAndres Camilo Buelvas MadariagaNo ratings yet
- Solucionario M. Cuantica Vol1Document227 pagesSolucionario M. Cuantica Vol1Luis Vargas Sanchez100% (1)
- Tarea CalorimetriaDocument4 pagesTarea CalorimetriaAlan HerreraNo ratings yet
- Predicción GeopresionesDocument26 pagesPredicción GeopresionesEso Payaso80% (5)
- MFI2670XEM Manual RefrigeradorDocument18 pagesMFI2670XEM Manual RefrigeradorJesus MoralesNo ratings yet