You might also like
- Dmdi U2 EaDocument9 pagesDmdi U2 Eavarzc13No ratings yet
- Problemas Resueltos de Balances de Materia y Energia PDFDocument30 pagesProblemas Resueltos de Balances de Materia y Energia PDFleyther2357% (7)
- Plan de negocios para una heladeríaDocument75 pagesPlan de negocios para una heladeríaBoris Castillo EspinosaNo ratings yet
- Plan de negocios para una heladeríaDocument75 pagesPlan de negocios para una heladeríaBoris Castillo EspinosaNo ratings yet
- NúcleoDocument11 pagesNúcleoGUILLERMO MURGA MASNo ratings yet
- Replicación de ADNDocument4 pagesReplicación de ADNNayeli HernándezNo ratings yet
- Clase Práctica No. 17 Cap. III. EnzimasDocument10 pagesClase Práctica No. 17 Cap. III. EnzimasAiden PalafoxNo ratings yet
- Formato de Planeación Didáctica UEMSTISDocument9 pagesFormato de Planeación Didáctica UEMSTISAlejandro Ferez HernandezNo ratings yet
- Biometría: Introducción a la estadística para las ciencias biológicasDocument175 pagesBiometría: Introducción a la estadística para las ciencias biológicashugo chungaNo ratings yet
- Acústica y ÓpticaDocument14 pagesAcústica y ÓpticaDibni CastilloNo ratings yet
- Ejemplo Diagrama de EstadosDocument4 pagesEjemplo Diagrama de EstadosJuan Carlos Pacheco GonzalezNo ratings yet
- Examen Final de Ciencias IIIDocument11 pagesExamen Final de Ciencias IIIAdelaida GonzálezNo ratings yet
- Cuestionario Geo Página 65Document1 pageCuestionario Geo Página 65Lorena Patricia AvilaNo ratings yet
- Óptica geométrica e instrumentos ópticosDocument22 pagesÓptica geométrica e instrumentos ópticosIsnardo BuenoNo ratings yet
- Calculo Vectorial 1.5 Al 1.7Document42 pagesCalculo Vectorial 1.5 Al 1.7adelangel5643No ratings yet
- RtosDocument56 pagesRtosElena_Huerta_H_1569No ratings yet
- Modelo 3D y cálculo de parámetros acústicos de una sala de teatroDocument8 pagesModelo 3D y cálculo de parámetros acústicos de una sala de teatroAndrey Santana AguileraNo ratings yet
- TtrtrytryttyDocument4 pagesTtrtrytryttypedroNo ratings yet
- Arbol Binario - 2193077915Document7 pagesArbol Binario - 2193077915Ricardo Josué Dominguez TorresNo ratings yet
- Instalación Apache Tomcat Windows guíaDocument14 pagesInstalación Apache Tomcat Windows guíaIvan Mac Gregor OviedoNo ratings yet
- Diagrama de Flujo 1Document2 pagesDiagrama de Flujo 1luis-o_tNo ratings yet
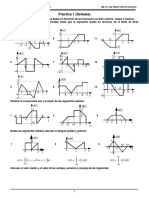
- Practica 1 Señales 2 2015Document4 pagesPractica 1 Señales 2 2015julian aruquipaNo ratings yet
- Diagrama de Flujo de DatosDocument5 pagesDiagrama de Flujo de DatosDariana MoralesNo ratings yet
- MICROOPROCESADORDocument7 pagesMICROOPROCESADORKarla QuintanaNo ratings yet
- Angel Alfredo Martinez BarrientosDocument19 pagesAngel Alfredo Martinez BarrientosAngelito AlfreditoNo ratings yet
- 7Document2 pages7Jason Porras PomaNo ratings yet
- (Pauta EDP) PDFDocument16 pages(Pauta EDP) PDFFelipe VergaraNo ratings yet
- Proyecto Autopartes-Cpml SasDocument89 pagesProyecto Autopartes-Cpml SasPaola Andrea ROMERO CERINZANo ratings yet
- Propuesta Diseño Red LanDocument11 pagesPropuesta Diseño Red LanLiliana AngelicaNo ratings yet
- Ecuaciones Diferenciales Parciales: Clasificación y Métodos de SoluciónDocument4 pagesEcuaciones Diferenciales Parciales: Clasificación y Métodos de SoluciónAndres WedemeyerNo ratings yet
- Diseño e implementación de cableado estructurado en instituto educativo de CuscoDocument12 pagesDiseño e implementación de cableado estructurado en instituto educativo de CuscoEmperatriz Sivana MarquezNo ratings yet
- Configuración básica de dispositivo de CiscoDocument3 pagesConfiguración básica de dispositivo de Ciscojsph123No ratings yet
- Edp QuicañoDocument175 pagesEdp QuicañoDAVID BRAYAN SALVATIERRA HUAMANINo ratings yet
- Reporte Practica 8Document6 pagesReporte Practica 8miriam arriagaNo ratings yet
- BBM1 U1 A2 VabcDocument3 pagesBBM1 U1 A2 VabcVane SchatzNo ratings yet
- DasdsadasdasdasdDocument48 pagesDasdsadasdasdasdJunior Sucasaca100% (1)
- Propuesta de Integración de TIDocument27 pagesPropuesta de Integración de TIDenia Isalia Santana Anguiano100% (1)
- Introduccion A La Fisica de La Guitarra ElectricaDocument2 pagesIntroduccion A La Fisica de La Guitarra ElectricaAlejandro Diaz Diaz0% (1)
- Tema 4 Componentes CelularesDocument33 pagesTema 4 Componentes Celularessofia villarmosaNo ratings yet
- Ejercicios de Programación EnsambladorDocument25 pagesEjercicios de Programación EnsambladorAída GabrielaNo ratings yet
- Función recursiva cuenta hojas árbol binarioDocument5 pagesFunción recursiva cuenta hojas árbol binarioMariana RodriguezNo ratings yet
- Representación Gráfica de SeñalesDocument12 pagesRepresentación Gráfica de SeñalesErick MendozaNo ratings yet
- CCNA2 Lab 11 2 1b EsDocument4 pagesCCNA2 Lab 11 2 1b Eskaryco4No ratings yet
- P4ABB D Santos RuizDocument4 pagesP4ABB D Santos RuizLizzete Santos RuizNo ratings yet
- Calcular probabilidad lluvia hierba húmeda red BayesianaDocument2 pagesCalcular probabilidad lluvia hierba húmeda red BayesianaRonald Alexander Valencia ValenciaNo ratings yet
- Componentes y aplicaciones de los sistemas expertosDocument8 pagesComponentes y aplicaciones de los sistemas expertosclaraNo ratings yet
- Interrupciones Del ProcesadorDocument3 pagesInterrupciones Del Procesadorwisin2010No ratings yet
- Arquitectura de OrdenadoresDocument17 pagesArquitectura de OrdenadoresWalter EduardoNo ratings yet
- Sistemas Operativos Paginacion y SegmentacionDocument71 pagesSistemas Operativos Paginacion y SegmentacionDavid Lopez FloresNo ratings yet
- Corrida financiera para refaccionaria de autosDocument37 pagesCorrida financiera para refaccionaria de autosJuan Carlos López Pardo100% (1)
- CD 2595Document3 pagesCD 2595Luis LoayzaNo ratings yet
- Pract10PrologOct2011.Doc 0Document6 pagesPract10PrologOct2011.Doc 0Javier SánchezNo ratings yet
- Practico Sistemas de Tempo RealDocument4 pagesPractico Sistemas de Tempo RealGuadalupe NassifNo ratings yet
- Niveles de RaidDocument13 pagesNiveles de RaidCasecNo ratings yet
- Planificadores de Sistemas de Tiempo RealDocument3 pagesPlanificadores de Sistemas de Tiempo Realpmaiuto2213No ratings yet
- Enzimas cinética inhibiciónDocument4 pagesEnzimas cinética inhibiciónAlex RolleroNo ratings yet
- BPRG U1 A3 CehrDocument3 pagesBPRG U1 A3 CehrvirhelgueraNo ratings yet
- Ejercicios de Probabilidades1Document3 pagesEjercicios de Probabilidades1Fernando JordanNo ratings yet
- Cuestionario de TecnologíaDocument2 pagesCuestionario de Tecnologíatecnologias999No ratings yet
- Tareabasesdedatos 1Document4 pagesTareabasesdedatos 1Gustavo López FigueroaNo ratings yet
- Objetivo 3. Transporte CelularDocument7 pagesObjetivo 3. Transporte Celularmusicart30008394100% (2)
- Funciones de la membrana celularDocument3 pagesFunciones de la membrana celularJavier Luna-PizarroNo ratings yet
- Guia Naturales Wilfrido ZabaletaDocument7 pagesGuia Naturales Wilfrido ZabaletaGeiner Luis Guzmán CorbachoNo ratings yet
- Crisis Alimenticia MexicoDocument10 pagesCrisis Alimenticia MexicoJohnNo ratings yet
- Balance de MateriaDocument22 pagesBalance de MateriaCarlos Sarmiento75% (8)
- Laboratorio de Bromatología de Forrajes Centro de Investigación en Nutrición AnimalDocument7 pagesLaboratorio de Bromatología de Forrajes Centro de Investigación en Nutrición AnimalErika Lopez ArandaNo ratings yet
- Laboratorio de Bromatología de Forrajes Centro de Investigación en Nutrición AnimalDocument7 pagesLaboratorio de Bromatología de Forrajes Centro de Investigación en Nutrición AnimalErika Lopez ArandaNo ratings yet
- Diagramas de Flujo y SimbologiaDocument14 pagesDiagramas de Flujo y SimbologiaNoe GallardoNo ratings yet
- Propiedades Termofísicas de AlimentosDocument38 pagesPropiedades Termofísicas de AlimentosRené MartínezNo ratings yet
- Conservación de AlimentosDocument22 pagesConservación de AlimentosFederico LeonNo ratings yet
- Aplicaciones de La Biotecnología PDFDocument124 pagesAplicaciones de La Biotecnología PDFLina Marcela Porras SamboniNo ratings yet
- Red de valor del mangoDocument8 pagesRed de valor del mangoShannon Gonzales VasquezNo ratings yet
- Revista Mexicana de Ciencias Agrícolas 2007-0934: IssnDocument8 pagesRevista Mexicana de Ciencias Agrícolas 2007-0934: IssnErika Lopez ArandaNo ratings yet
- Seminario ExtraccionDocument24 pagesSeminario ExtraccionErika Lopez ArandaNo ratings yet
- Revista de Simulacion y Laboratorio V3 N9 4Document8 pagesRevista de Simulacion y Laboratorio V3 N9 4JonattanSaulNo ratings yet
- Apuntes Balance de Materia y Energia 2004Document88 pagesApuntes Balance de Materia y Energia 2004Jose Kalvin Rojas Sosa89% (9)
- Revista Iberoamericana de Tecnología Postcosecha 1665-0204: IssnDocument9 pagesRevista Iberoamericana de Tecnología Postcosecha 1665-0204: IssnErika Lopez ArandaNo ratings yet
- Consumo Saludable CiDocument44 pagesConsumo Saludable CiErika Lopez Aranda100% (1)
- Deshidratacion Osmotica en Palta Variedad FuerteDocument79 pagesDeshidratacion Osmotica en Palta Variedad Fuerteroosvelt vasquez chicomaNo ratings yet
- 1424895611tendencias RU Alimentos Funcionales 2014Document27 pages1424895611tendencias RU Alimentos Funcionales 2014Erika Lopez ArandaNo ratings yet
- Monografia 6Document6 pagesMonografia 6Yuuki Sakura KuranNo ratings yet
- Alimentacion Del Ganado CaprinoDocument22 pagesAlimentacion Del Ganado CaprinoPaola Serrano DazaNo ratings yet
- Camara Frigorifica Funcionamiento PDFDocument53 pagesCamara Frigorifica Funcionamiento PDFdoritaxx0% (1)
- Cuadro Con La Taxonomía de Bloom 03Document5 pagesCuadro Con La Taxonomía de Bloom 03mauriciorvillalobos5993No ratings yet
- 11 CarbohidratosDocument77 pages11 CarbohidratosMario Leonardo Guevara EscalanteNo ratings yet
- Unidad II SecadoDocument5 pagesUnidad II SecadoErika Lopez ArandaNo ratings yet
- 003 Estructura Molecular Parte 5 5875Document11 pages003 Estructura Molecular Parte 5 5875Erika Lopez ArandaNo ratings yet
- Columna de Extraccion Liquido LiquidoDocument44 pagesColumna de Extraccion Liquido LiquidoLeon LuisNo ratings yet
- Transferencia Molecular de Calor, Masa y Cantidad de MovimientoDocument424 pagesTransferencia Molecular de Calor, Masa y Cantidad de MovimientoHugo Cruz Teles100% (5)
- Columna de Extraccion Liquido LiquidoDocument44 pagesColumna de Extraccion Liquido LiquidoLeon LuisNo ratings yet
- Preparación de soluciones conservantes para frutas agroexportaciónDocument71 pagesPreparación de soluciones conservantes para frutas agroexportaciónGARY FERNANDO GARCÍA LEÓNNo ratings yet
- Transporte de Cargas en Sistemas BiologicosDocument2 pagesTransporte de Cargas en Sistemas Biologicosyoemis100% (1)
- Evaluación Diagnóstica de Ciencia Y Tecnología: ¿Por Qué Se Oscurecen Algunos AlimentosDocument3 pagesEvaluación Diagnóstica de Ciencia Y Tecnología: ¿Por Qué Se Oscurecen Algunos AlimentosJose Carlos Quispe espinozaNo ratings yet
- TRANSFORMACIÓN AERÓBICA - ResumenDocument5 pagesTRANSFORMACIÓN AERÓBICA - ResumenJuan Jeferson AlosillaNo ratings yet
- Determinación de GlucosaDocument4 pagesDeterminación de GlucosaglodovichiNo ratings yet
- Maduracion Frutos PDFDocument11 pagesMaduracion Frutos PDFEdith Raquel Macias GarciaNo ratings yet
- Estructura Jerarquica de Las Proteinas (LODISH 5ta) PDFDocument8 pagesEstructura Jerarquica de Las Proteinas (LODISH 5ta) PDFvalenromeroNo ratings yet
- Practica N°7 Oxidaciones Biologicas. LVDocument6 pagesPractica N°7 Oxidaciones Biologicas. LVluianvil3No ratings yet
- Bioquimica Enzimas 2Document13 pagesBioquimica Enzimas 2Eli Motta AlcocerNo ratings yet
- Apo #7 - EnzimasDocument53 pagesApo #7 - Enzimasanapokemon0072No ratings yet
- .Metabolismo de AminoacidosDocument9 pages.Metabolismo de AminoacidosFlorencia LahozNo ratings yet
- Existen Enzimas SintéticasDocument3 pagesExisten Enzimas SintéticasNatalyGallosoSanchezNo ratings yet
- Glucosa en orina y sangreDocument23 pagesGlucosa en orina y sangreJisila JisilaNo ratings yet
- Bioquimica Informe. para Revisar y CompletarDocument13 pagesBioquimica Informe. para Revisar y CompletarYelina CarrascoNo ratings yet
- Práctica #7 InmovilizaciónDocument5 pagesPráctica #7 InmovilizaciónSandra MedinaNo ratings yet
- Problem A RioDocument10 pagesProblem A RioItseel EONo ratings yet
- Control biológico de Botrytis cinerea en uva con Clonostachys roseaDocument8 pagesControl biológico de Botrytis cinerea en uva con Clonostachys roseaJuan Pablo Vásquez LozadaNo ratings yet
- Cinética ExposiciónDocument106 pagesCinética ExposiciónYanella Smith Arevalo GonzalesNo ratings yet
- Minerales Alimentacion AnimalDocument149 pagesMinerales Alimentacion AnimalchatrumanNo ratings yet
- Inmovilización de EnzimasDocument7 pagesInmovilización de EnzimasMarcos GalárragaNo ratings yet
- Enzimas en ClínicaDocument45 pagesEnzimas en ClínicaLilian ArchivosNo ratings yet
- ENZIMASDocument22 pagesENZIMASWendi Carolina Santos HernandezNo ratings yet
- Informe EnzimasDocument14 pagesInforme Enzimasdaniso12No ratings yet
- Elaboración de cerveza de trigoDocument45 pagesElaboración de cerveza de trigocarmen rosa floresNo ratings yet
- Proceso Productivo de La Elaboración de Cerveza Lager A Escala IndustrialDocument110 pagesProceso Productivo de La Elaboración de Cerveza Lager A Escala IndustrialhgyjkllNo ratings yet
- Causas por las que se descompone un alimentoDocument3 pagesCausas por las que se descompone un alimentoJOCELYN MARIN100% (1)
- BioquímicaDocument9 pagesBioquímicaFernanda NuñezNo ratings yet
- Grupo 3. Taller Extracción ADN Candida.Document3 pagesGrupo 3. Taller Extracción ADN Candida.Diego CalderaNo ratings yet
- Amilasa SalivalDocument28 pagesAmilasa SalivalCarlos Mario AtenciaNo ratings yet
- Metodos de Conservacion y Quimica de Los AlimentosDocument105 pagesMetodos de Conservacion y Quimica de Los AlimentosmalorieNo ratings yet