You might also like
- Wecb657 PDFDocument10 pagesWecb657 PDFazzaassNo ratings yet
- Wecb657 PDFDocument10 pagesWecb657 PDFazzaassNo ratings yet
- Wecb 467Document31 pagesWecb 467qhqhqNo ratings yet
- Wecb439 PDFDocument12 pagesWecb439 PDFazzaassNo ratings yet
- Wecb641 PDFDocument15 pagesWecb641 PDFazzaassNo ratings yet
- Wecb451 PDFDocument7 pagesWecb451 PDFazzaassNo ratings yet
- Wecb451 PDFDocument7 pagesWecb451 PDFazzaassNo ratings yet
- Wecb641 PDFDocument15 pagesWecb641 PDFazzaassNo ratings yet
- The Pentose Phosphate Pathway: An Overview: Advanced ArticleDocument12 pagesThe Pentose Phosphate Pathway: An Overview: Advanced ArticleazzaassNo ratings yet
- Wecb434 PDFDocument13 pagesWecb434 PDFazzaassNo ratings yet
- Wecb 431Document12 pagesWecb 431azzaassNo ratings yet
- Wecb435 PDFDocument10 pagesWecb435 PDFazzaassNo ratings yet
- Wecb 424Document12 pagesWecb 424azzaassNo ratings yet
- Wecb651 PDFDocument15 pagesWecb651 PDFazzaassNo ratings yet
- Wecb434 PDFDocument13 pagesWecb434 PDFazzaassNo ratings yet
- Passive Diffusion Across Membranes: Advanced ArticleDocument10 pagesPassive Diffusion Across Membranes: Advanced ArticleazzaassNo ratings yet
- Wecb 423Document7 pagesWecb 423azzaassNo ratings yet
- Wecb 417Document10 pagesWecb 417azzaassNo ratings yet
- The Pentose Phosphate Pathway: An Overview: Advanced ArticleDocument12 pagesThe Pentose Phosphate Pathway: An Overview: Advanced ArticleazzaassNo ratings yet
- Passive Diffusion Across Membranes: Advanced ArticleDocument10 pagesPassive Diffusion Across Membranes: Advanced ArticleazzaassNo ratings yet
- Wecb 527Document11 pagesWecb 527azzaassNo ratings yet
- Wecb 431Document12 pagesWecb 431azzaassNo ratings yet
- Wecb 424Document12 pagesWecb 424azzaassNo ratings yet
- Wecb 424Document12 pagesWecb 424azzaassNo ratings yet
- Wecb 431Document12 pagesWecb 431azzaassNo ratings yet
- Wecb 424Document12 pagesWecb 424azzaassNo ratings yet
- Wecb 688Document14 pagesWecb 688azzaassNo ratings yet
- Wecb 527Document11 pagesWecb 527azzaassNo ratings yet
- Wecb 423Document7 pagesWecb 423azzaassNo ratings yet
- Wecb 417Document10 pagesWecb 417azzaassNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5784)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- 03 Modul SC T2-Bab3 (Csy4p) PDFDocument22 pages03 Modul SC T2-Bab3 (Csy4p) PDFSitiRohaizaMatYusoffNo ratings yet
- Drug Study Ferrous SulfateDocument2 pagesDrug Study Ferrous SulfateBunnie AlphaNo ratings yet
- HemoglobinopathiesDocument26 pagesHemoglobinopathiesSanjey GaneshNo ratings yet
- MUST KNOW HEMATOLOGYDocument44 pagesMUST KNOW HEMATOLOGYJan Joseph YbañezNo ratings yet
- Hariom Kumar RoyDocument24 pagesHariom Kumar RoyRajeev Kumar Roy0% (1)
- IBO 2018 - Theoretical Exam ADocument51 pagesIBO 2018 - Theoretical Exam ALelouch vi Britannia The 3rd100% (1)
- (Bilirubin-Albumin) Binding and Neonatal JaundiceDocument6 pages(Bilirubin-Albumin) Binding and Neonatal JaundiceErwin SmithNo ratings yet
- 1B.2 Transport of Oxygen and Fetal HeamoglobinDocument35 pages1B.2 Transport of Oxygen and Fetal Heamoglobintas xoNo ratings yet
- What Is The Complete Blood Count Test (CBC) ?: White Blood Cells Red Blood Cells PlateletsDocument4 pagesWhat Is The Complete Blood Count Test (CBC) ?: White Blood Cells Red Blood Cells PlateletsSheryl Ann Celis BacaniNo ratings yet
- Surgery Examination - 3-4 (Question Cases) (Eng)Document45 pagesSurgery Examination - 3-4 (Question Cases) (Eng)Aiman TymerNo ratings yet
- Oxyganation: Is IsDocument5 pagesOxyganation: Is IsRahmatun NurainaNo ratings yet
- Chapter 2 Respiration Sec 3Document11 pagesChapter 2 Respiration Sec 3Qian hui TanNo ratings yet
- AEEE 2024 Sample PaperDocument98 pagesAEEE 2024 Sample Paperaiswaryasakthivel007No ratings yet
- Report HB ELECTROPHORESISDocument2 pagesReport HB ELECTROPHORESISShweta PatilNo ratings yet
- Hemoglobin and Myoglobin Structure, Function and GeneticsDocument48 pagesHemoglobin and Myoglobin Structure, Function and GeneticsLuis VicenteNo ratings yet
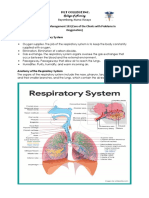
- Respiratory System Anatomy, Assessment & Diagnostic TestsDocument24 pagesRespiratory System Anatomy, Assessment & Diagnostic TestsPrince Rener Velasco PeraNo ratings yet
- ThalassemiaDocument24 pagesThalassemiaafif100% (1)
- Diagnostic Pitfalls of Less Well Recognized HBHDocument10 pagesDiagnostic Pitfalls of Less Well Recognized HBHLink BuiNo ratings yet
- Lec - M4. Post Task I - Cardiovascular SystemDocument2 pagesLec - M4. Post Task I - Cardiovascular SystemEmily BernatNo ratings yet
- QigongBreath PDFDocument12 pagesQigongBreath PDFfofofofo100% (2)
- Bio F110 1001Document2 pagesBio F110 1001AmanNo ratings yet
- ASCP Questions (Autosaved) - 1Document56 pagesASCP Questions (Autosaved) - 1Glenn Perez100% (16)
- Physiology, Oxygen Transport: April 2019Document5 pagesPhysiology, Oxygen Transport: April 2019Soha SonaNo ratings yet
- 36 ch17 PDFDocument19 pages36 ch17 PDFShivamNo ratings yet
- HEMOGLOBINOPATHY GUIDEDocument3 pagesHEMOGLOBINOPATHY GUIDEChatie PipitNo ratings yet
- Oxygen and Carbon Dioxide Transport in the Respiratory SystemDocument108 pagesOxygen and Carbon Dioxide Transport in the Respiratory SystemrifqifuadiNo ratings yet
- CAIE Biology A-level Transport in Mammals FlashcardsDocument77 pagesCAIE Biology A-level Transport in Mammals FlashcardsElisaNo ratings yet
- Trio Service Manual DomesticDocument81 pagesTrio Service Manual DomesticGiovanny RamirezNo ratings yet
- 2010 Teachers GuideDocument90 pages2010 Teachers GuidebiswacementNo ratings yet
- Dr. Kamal KV FINAL BOOST PDFDocument41 pagesDr. Kamal KV FINAL BOOST PDFReeba janetNo ratings yet