!"#$"% '( )**+,-+".
/ 0112 3 4*5 )%(6"#'*7 8"6$*+
"
I. Anatoma y Fisiologa del Sistema Excito-Conductor
A. Anatoma del Sistema Excito-Conductor
El sistema excito-conductor del corazon comprende un conjunto de celulas especializadas
que inician y transmiten la actividad electrica responsable de las contracciones
coordinadas de las camaras cardiacas.
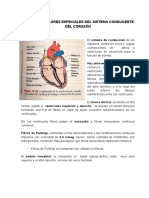
El ndulo sinusal, constituido por una pequea masa de celulas miocardicas
especializadas, ubicadas a la derecha de la desembocadura de la vena cava superior, inicia
normalmente el impulso electrico del corazon.
El ndulo aurculo-ventricular yace bajo el endocardio en la region inIero-posterior del
septum interauricular. Distal al nodo auriculo-ventricular se encuentra el haz de His el
que perIora hacia posterior el septum interventricular. Dentro del septum el haz de His se
biIurca en a) una gruesa sabana de Iibras que se continua hacia el borde izquierdo del
septum constituyendo la rama izquierda del has de His y en b) una estructura compacta
en Iorma de cable hacia la derecha denominada rama derecha del haz de His.
La rama derecha es gruesa y se encuentra inmersa en la porcion muscular del septum
interventricular. De ahi se continua hacia el apex biIurcandose en el punto de la union
del septum y la pared anterior del ventriculo derecho. A ese nivel la rama derecha se
localiza en la region subendocardica. Una de las ramas biIurcadas se continua a traves de
la banda moderadora mientras que la otra se dirige hacia la punta. Ambas ramas se
subdividen constituyendo un plexo a lo largo y ancho de la camara ventricular.
La rama izquierda, desde un punto de vista Iuncional se divide en un fascculo anterior y
otro posterior. El Iasciculo anterior recorre la cara anterior del ventriculo izquierdo hacia
el apex Iormando un plexo subendocardico en relacion al musculo papilar anterior,
mientras que el posterior se dirige hacia el musculo papilar posterior para luego constituir
tambien un plexo subendocardico en el resto del ventriculo izquierdo.
Los plexos subendocardicos de ambos ventriculos distribuyen fibras de Purkinje al
miocardio ventricular. Los impulsos provenientes del sistema His-Purkinje se transmiten
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
#
a los musculos papilares y en seguida a las paredes ventriculares. Esto ultimo trae como
consecuencia el que la contraccion de los musculos papilares preceda a la del resto de los
ventriculos lo que evita la regurgitacion sistolica de sangre a nivel de las valvulas
auriculo-ventriculares.
Un esquema de la anatomia del sistema excito conductor se representa en la Iigura 1
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
$
B. Inervacin del Corazn] XE "Inervacin del Corazn" }
El corazn recibe inervacin simptica y parasimptica. Las neuronas
preganglionares simpticas se localizan en los primeros segmentos torcicos de la
mdula espinal y sinaptan con neuronas de segundo orden ubicadas en los ganglios
simpticos cervicales. En un recorrido al interior de los nervios cardiacos se puede
observar que estas fibras llegan al corazn y los grandes vasos. Las fibras
preganglionares parasimpticas se originan en el ncleo motor de la medula y pasan
como ramos del nervio vago al corazn y grandes vasos. A este nivel las fibras
sinaptan con neuronas de segundo orden localizadas en ganglios intratorcicos.
Fibras aferentes vagales ubicadas en la pared inferior y posterior de los ventriculos
dan origen a importantes reflejos cardiacos, mientras que eferencias vagales a nivel del
ndulo sinusal y auriculo-ventricular juegan un rol muy significativo en la
modulacin de los impulsos elctricos.
C. Electrofisiologa Bsica
a- Potencial de Reposo
Las Iibras cardiacas en reposo se encuentran polarizadas, vale decir, exhiben una
diIerencia de potencial entre el medio intracelular y el extracelular siendo, el interior
negativo respecto al exterior (Iig 2). Esta diIerencia de potencial durante la diastole
electrica se denomina potencial de reposo transmembrana (PRT) y su valor depende del
tipo de Iibra (-90 mV para las Iibras auriculares, ventriculares y del sistema His Purkinje;
-60 mV para Iibras del nodulo sinusal y nodo auriculoventricular). El PRT es estable en
las Iibras no automaticas; en aquellas dotadas de automatismo se produce una
depolarizacion diastolica, la que es mas pronunciada en las celulas del nodulo sinusal.
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
%
Fig 2. DiIerencia de potencial entre el medio intracelular y extracelular
registrada por microelectrodos en una Iibra cardiaca durante la
diastole electrica. PRT Potencial de reposo transmembrana
b- Potencial de Accion
Este traduce variaciones del potencial transmembrana en Iuncion del tiempo. El potencial
de accion esta compuesto por varias Iases. La Iase ascendente del potencial de accion se
denomina Iase 0 y corresponde a la depolarizacion de la celula. La repolarizacion inicial
se denomina Iase 1, la que se continua con un plateau o Iase 2. La Iase 3 (repolarizacion)
lleva el potencial transmembrana a los niveles de reposo. La Iase 4 corresponde a la Iase
de reposo diastolico.(Iig 3 )
Fig 3. Fases del Potencial de accion (PA) PRT: potencial de reposo transmembrana;
PU: potencial umbral; mV: milivoltios T: tiempo en milisegundos
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
&
c- Potencial Umbral
Corresponde al valor del potencial transmembrana a partir del cual se genera un potencial
de accion. En las Iibras no automaticas es alcanzado por Ilujos electrotonicos que
proceden de Iibras vecinas depolarizadas. En las celulas automaticas el potencial umbral
puede alcanzarse por la depolarizacion diastolica espontanea de sus Iibras.
d- Tipos de Potencial de Accion
Existen Iundamentalmente dos tipos de potencial de accion:
a) Potencial de accion de Iibras rapidas (auriculas, ventriculos, His Purkinje). Se
caracteriza por una Iase 0 de inscripcion rapida y de gran amplitud. La alta
velocidad de ascenso de la Iase 0 determina que en estos tejidos la velocidad de
conduccion sea elevada (0.5-5 m/sec).
b) Potencial de accion de Iibras lentas (nodulo sinusal y nodulo auriculo-
ventricular). En ellas la Iase 0 es de inscripcion lenta y amplitud disminuida. La
velocidad de conduccion de estas Iibras es de solo 0.01-0.1 m/sec.
La Iig 4 representa los 2 tipos de potencial de accion (A : Iibra rapida; B: Iibra lenta )
La Iigura 5 muestra los distintos tipos de potenciales de accion de las Iibras cardiacas y
su relacion con el ECG de superIicie.
El nivel del PRT de las Iibras cardiacas determina la velocidad de ascenso y la amplitud
del potencial de accion. A mayor negatividad del PRT mayor sera la velocidad de ascenso
y la amplitud de este.
En situaciones de anoxia, isquemia, aumento del potasio extracelular o por intoxicacion
digitalica, la membrana puede ser llevada a un estado de hipopolarizacion (menor
negatividad del PRT). A la inversa, en presencia de hipokalemia el PRT se hace mas
negativo lo que determina un aumento de la velocidad de ascenso de la Iase 0, un
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
'
aumento de la amplitud del potencial de accion y una mayor velocidad de conduccion.
Fig 4. Tipos de Potencial de accion. Arriba : Iibra rapida ; Abajo : Iibra lenta
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
(
Fig 5. Potenciales de accion cardiacos y su relacion con el ECG de superIicie
D. Aspectos inicos de la actividad elctrica del corazn
a- Potencial de reposo.
El estado de polarizacin de las fibras cardiacas en reposo se debe a la diferente
concentracin inica entre el medio intracelular y el extracelular.
Esta desigual reparticin de iones entre ambos medios es mantenida por un mecanismo
activo que consume energia. Para el sodio y el potasio funciona una bomba que
expulsa el sodio e incorpora el potasio a la clula. Esta bomba es activada por
magnesio y la energia que ocupa es aportada por la degradacin de A1P.
La bomba sodio-potasio funciona de manera asimtrica, de tal suerte que la corriente
sdica de salida es de mayor magnitud que la corriente de entrada potsica. Como
consecuencia de este funcionamiento asimtrico se genera el potencial de reposo
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
)
transmembrana. En cuanto a la salida de calcio, tambin intervendria una bomba que
utiliza energia proveniente de la degradacin del A1P. La salida del calcio depende de
la gradiente de concentracin de sodio y por consiguiente es influida por la bomba
sodio-potasio.
Las corrientes pasivas inicas transmembrana se rigen por la ley de Ohm:
I g (Vm - Ei)
I : Flujo inico. g: Conductancia. Vm:Potencial
transmembrana.
Ei: Potencial de
equilibrio.
A su vez la conductancia (g) representa el valor inverso de la resistencia al paso de un
determinado ion.
1
g -----
R
R: Resistencia.
b- Potencial de equilibrio.
Cada ion durante la distole elctrica estar sometido a dos fuerzas: una quimica dada
por el gradiente de concentracin (que tender a introducir sodio y calcio y a sacar
potasio de las clulas) y otra elctrica dada por gradiente de potencial (que tender a
introducir cationes al medio intracelular). Para cada ion existir un valor de potencial
transmembrana en el cual ambas fuerzas se anulan (la gradiente elctrica se equilibra
con la gradiente quimica). Alcanzado este valor de potencial, el flujo neto
transmembrana de dicho ion ser nulo.
El valor de potencial transmembrana en el cual un determinado ion tiene un flujo
igual a por anulacin de gradientes, se conoce con el nombre de potencial de
equilibrio, el que puede calcularse por la ecuacin de Aernst.
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
*
RT XE
Ex --------- In ---------
ZF Xi
Ex: Potencial de equilibrio para un determinado ion.
R : Constante del gas perfecto.
T : Tiempo absoluto.
Z : Valencia del ion.
F : Valor de Faraday : unidad de capacitancia.
In : Logaritmo natural.
XE: Concentracin extracelular del ion X.
X i: Concentracin intracelular del ion X.
Aplicando la ecuacin de Nernst para cada ion tenemos:
RT K+ EC
EK+ ------- In ------------ - 95 mv
ZF K+ I C
RT Na+EC
ENa+ --------- In ------------ + 60 mv
ZF Na+IC
RT Ca++ EC
ECa++ ------- In -------------- + 80 mv
ZF Ca++ IC
Vm - ENa+ - 150 (flujo de entrada)
Vm - EK+ + 5 (flujo de salida)
Vm-ECa++ -170 (flujo de entrada)
Si a pesar de que el valor Jm - EAa es 3 veces mayor que el valor Jm - EK ( y a pesar
de eso ambos flujos se equilibran) significa que la membrana en reposo es 3 veces
ms permeable al potasio que al sodio. (En las fibras lentas en que el PR1 es menor, la
permeabilidad al potasio es tambin menor).
El grado de polarizacin de la membrana en reposo depende de la relacin entre la
concentracin extracelular e intracelular de potasio. De acuerdo a la ecuacin de
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
"+
Aernst, todo aumento de potasio intracelular o disminucin de potasio extracelular
lleva a un estado de hiperpolarizacin, mientras que una disminucin del potasio
intracelular o un aumento del potasio extracelular determina una hipopolarizacin .
E. Corrientes inicas.
Las variaciones de potencial que ocurren durante el potencial de accion se deben al paso
de iones en uno y otro sentido a traves de la membrana celular. Dado que estos Ilujos
ocurren en el sentido de las gradientes ionicas no consumen energia (Ilujos pasivos).
Las corrientes ionicas pueden ser depolarizantes o repolarizantes. En el primer caso,
habra un Ilujo neto de cargas positivas al interior de las celulas; en el segundo habra una
salida de cargas positivas al medio extracelular.
Si la diIerencia entre el potencial de membrana y el potencial de equilibrio para un ion da
un valor positivo la corriente ionica sera repolarizante (es el caso del potasio); si la
diIerencia entre ambos valores es negativa la corriente sera depolarizante (es el caso del
sodio).
F. Corrientes inicas durante el potencial de accin.
1. Fibras rpidas. Fase 0: Esta Iase se debe a un aumento brusco y transitorio de la
conductancia sodica, lo que determina una corriente de entrada de dicho ion. Esto
ocurrira cuando el PRT sea llevado al valor de potencial umbral.
La corriente sdica de entrada depende de dos variables representadas por compuertas
ubicadas en serie a travs de las cuales el ion penetra al interior de las clulas.
Estas dos variables corresponden a una de activacin (compuerta m) y otra de
inactivacin (compuerta h).
La compuerta m est cerrada (m ) cuando el potencial de membrana es de -8 mv y
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
""
a partir de ese valor se comienza a abrir, llegando a un grado de apertura mxima (m
1) cuando el potencial de membrana es de 2 mv. La constante de tiempo de esta
variable de activacin es de 1 mseg lo que significa que las variaciones de
permeabilidad en funcin del tiempo para la entrada de sodio son muy rpidas. La
variable de inactivacin se representa por una compuerta h, la que se encuentra
abierta cuando el potencial transmembrana es de -8 mv (h 1) y comienza a cerrarse
con valores menores de potencial de membrana, llegando a ocluirse totalmente (h )
cuando el potencial de membrana llega a -55 mv. La compuerta h inicia
posteriormente una lenta apertura que culmina al final de la repolarizacin. La
constante de tiempo de la variable de inactivacin es algo ms larga que la
correspondiente a la variable de activacin.
El paso de sodio depende por una parte del mayor o menor grado de apertura de ambos
tipos de compuertas y por otra de la relacin existente en un momento dado, entre el
potencial de membrana y el potencial de equilibrio para el ion sodio.
El canal representado por las compuertas m y h se denomina canal rpido. Este es
inhibido especificamente por la tetrodotoxina. La corriente de entrada sdica rpida es
la responsable de la fase inicial de la depolarizacin de las fibras fuertemente
polarizadas en reposo.
En un rango de voltaje entre -5 y +2 mv la corriente hacia adentro depende de
variables distintas, tambin representadas como variables de activacin (compuerta d)
y de inactivacin (compuerta f). A travs de estas compuertas penetran iones calcio y
sodio. Otros cationes tambin permeables a este canal son el estroncio, el bario y el
magnesio. Las variables de activacin y de inactivacin representadas por las
compuertas d y f tienen constantes de tiempo mucho mayores. De ahi el nombre
asignado a esta corriente de entrada (canal lento). El canal lento es inhibido por
verapamilo y manganeso y es el responsable de la mantencin de la depolarizacin una
vez que el canal rpido se ha inactivado.
Fase 1. Esta Iase tambien llamada repolarizacion inicial estaria dada por una corriente de
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
"#
salida de K a traves de la activacion de un canal transitorio de dicho ion.
Fase 2 o plateau. En su genesis interviene la entrada de sodio y calcio a traves del canal
lento y una disminucion de la corriente de salida del ion potasio de base (llamada gK 1).
Esta Iase es visible especialmente en las Iibras ventriculares y del sistema His Purkinje.
Fase 3 o repolarizacion terminal. Se debe a la inactivacion del canal lento sodico-calcico
por una parte y a la aparicion de una corriente de salida de ion potasio ( llamada IX 1) por
otra. Al Iinal de la repolarizacion se produce ademas un aumento de la corriente de salida
de potasio de base (gK 1 ).
Fibras lentas. En ellas el potencial umbral es de alrededor de -50 mv. A estos niveles de
potencial, el canal sodico rapido esta inactivado en gran parte. La depolarizacion en las
Iibras lentas es dependiente del canal sodico-calcico lento.
La repolarizacion al igual que en las Iibras rapidas depende de la inactivacion del canal
lento por una parte y de la salida del ion potasio (IX 1).
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
"$
G. Excitabilidad de las fibras cardacas.
La excitabilidad de las Iibras cardiacas corresponde a la propiedad que poseen de generar
un potencial de accion como consecuencia de un estimulo aplicado a ellas.
Durante gran parte del potencial de accion, la Iibra no es excitable cualquiera sea la
intensidad de la estimulacion recibida. A esto se denomina Iase de reIractariedad.
El perodo refractario absoluto de la Iibra corresponde a un estado de inexcitabilidad
total. Este estado existe durante las Iases 0, 1, 2 y en parte de la Iase 3 en las Iibras
rapidas. A partir de un valor de potencial de membrana de -55 mv, la Iibra recupera
parcialmente su excitabilidad, lo que se demuestra estimulando la Iibra con corrientes
supraumbrales. Las primeras respuestas generadas no son propagadas.
El perodo refractario efectivo incluye el periodo reIractario absoluto y aquel con
respuestas generadas no propagadas. Este termina cuando aparecen las primeras
respuestas propagadas. Estas ultimas sobrevienen a partir de potenciales poco negativos y
por otra parte en momentos en que las conductancias de los canales de depolarizacion no
estan aun recuperadas. Debido a esto los potenciales de accion que pueden sobrevenir en
esta etapa tienen Iase 0 lenta y de escasa amplitud.
El perodo refractario relativo incluye el periodo reIractario absoluto, el eIectivo y
aquel en el cual las Iibras solo son excitables con corrientes supraumbrales.
Al final de la fase 3, en las fibras rpidas puede aparecer un periodo corto en que
estimulos subumbrales son capaces de generar un potencial de accin (fase
supernormal de excitabilidad). Este mayor grado de excitabilidad de las fibras, sin
embargo, ocurre durante potenciales de membrana poco negativos, y por tanto, las
respuestas generadas son lentas. Por otra parte, el grado de recuperacin de la
excitabilidad en este periodo es heterogneo, lo que conlleva el riesgo de respuestas
desincronizadas a partir de un estimulo nico (fase vulnerable).
El periodo reIractario total termina con el retorno de la excitabilidad completa despues de
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
"%
la Iase supernormal.
Se denomina perodo refractario funcional de la Iibra al intervalo mas corto que separa
2 respuestas normalmente propagadas.
Los periodos reIractarios que deIinen las Iases de la excitabilidad cardiaca son
esquematizados en la Iigura 6
Fig 6. Periodos reIractarios de las Iibras cardiacas
En las fibras lentas, el periodo refractario efectivo va mas alla de la duracion del
potencial de accion. Esto se debe a la mavor constante de tiempo del canal lento (en
comparacion con el canal rapido) lo que hace que este no este en condiciones de
reactivarse a pesar de haberse completado el potencial de accion. La duracion de los
potenciales de accion v de los periodos refractarios es funcion de la longitud del ciclo
cardiaco precedente (a mavor longitud del ciclo precedente mavor duracion del
potencial de accion v de los periodos refractarios). Esto explica la aparicion de
aberrancias ventriculares por un latido anticipado cuando este ha sido precedido por un
ciclo largo (fenomeno de Ashman).
!"#$"% '( )**+,-+"./ 0112 3 4*5 )%(6"#'*7 8"6$*+
"&
H. Automatismo Cardiaco
Esta propiedad de algunas Iibras cardiacas se debe a la presencia en ellas de una
depolarizacion diastolica espontanea. La pendiente de depolarizacion diastolica es mas
pronunciada en el nodulo sinusal, que en otras Iibras automaticas.
Esta depolarizacion espontanea durante la diastole se debe a un Ilujo de entrada de iones
denominado If. Las evidencias actuales involucran predominantemente al Na en este
Ienomeno el que penetra por canales diIerentes de los canales de Na rapidos descritos
anteriormente.
I. Conduccin Cardiaca
Esta se eIectua desde las Iibras depolarizadas a las Iibras vecinas polarizadas a traves de
conecciones de baja resistencia al paso de los impulsos ('gap junctions) Corresponde a
un Ienomeno electrico (transmision electrotonica) dado por la diIerencia de potencial que
se produce entre las celulas activadas y las celulas en reposo, lo que condiciona un Ilujo
de corriente. La velocidad de conduccion es dependiente del grosor de las Iibras cardiacas
(a mayor grosor mayor velocidad de conduccion) del tipo de union intercelular y de la
disposicion geometrica de las Iibras (la convergencia de Iibras mejora la conduccion; la
divergencia la empeora).
Este libro fue distribuido por cortesa de:
Para obtener tu propio acceso a lecturas y libros electrnicos ilimitados GRATIS
hoy mismo, visita:
http://espanol.Free-eBooks.net
Comparte este libro con todos y cada uno de tus amigos de forma automtica,
mediante la seleccin de cualquiera de las opciones de abajo:
Para mostrar tu agradecimiento al autor y ayudar a otros para tener
agradables experiencias de lectura y encontrar informacin valiosa,
estaremos muy agradecidos si
"publicas un comentario para este libro aqu".
INFORMACIN DE LOS DERECHOS DEL AUTOR
Free-eBooks.net respeta la propiedad intelectual de otros. Cuando los propietarios de los derechos de un libro envan su trabajo a Free-eBooks.net, nos estn dando permiso para distribuir dicho
material. A menos que se indique lo contrario en este libro, este permiso no se transmite a los dems. Por lo tanto, la redistribucin de este libro sn el permiso del propietario de los derechos, puede
constituir una infraccin a las leyes de propiedad intelectual. Si usted cree que su trabajo se ha utilizado de una manera que constituya una violacin a los derechos de autor, por favor, siga nuestras
Recomendaciones y Procedimiento de Reclamos de Violacin a Derechos de Autor como se ve en nuestras Condiciones de Servicio aqu:
http://espanol.free-ebooks.net/tos.html
You might also like
- Nueva Medicina Germánica (Parte I) - Dr. Ryke Geerd HamerDocument576 pagesNueva Medicina Germánica (Parte I) - Dr. Ryke Geerd Hamereoeoe69100% (90)
- ReflexologiaDocument60 pagesReflexologiaskario8100% (1)
- Primer Manual de Nutirición ConcienteDocument333 pagesPrimer Manual de Nutirición ConcienteJosé María Morales98% (42)
- El Cerebro Del CorazónDocument6 pagesEl Cerebro Del Corazónrosica1973No ratings yet
- La Nueva Medicina Germánica Del DR HamerDocument34 pagesLa Nueva Medicina Germánica Del DR Hameryaco63100% (2)
- Geometría Sagrada Imprimir PDFDocument87 pagesGeometría Sagrada Imprimir PDFlordkur100% (4)
- Aura MasterDocument65 pagesAura MasterJose Luis Ferrario P100% (2)
- Dietoterapia Energética - Patricia GuerinDocument227 pagesDietoterapia Energética - Patricia GuerinLittleSpam95% (22)
- Guia Completa para El Ayuno PDFDocument17 pagesGuia Completa para El Ayuno PDFrosica1973100% (1)
- La Dieta Alcalina PDFDocument10 pagesLa Dieta Alcalina PDFDavid CaridadNo ratings yet
- El Libro Del ShiatsuDocument191 pagesEl Libro Del ShiatsuRy Ortega0% (1)
- Buenas Practicas de OrdeñoDocument9 pagesBuenas Practicas de OrdeñoRaul Hinostroza TenorioNo ratings yet
- Comer y No Morir en El Intento-7 Ed - Mayo09-Aditivos, Cosmeticos, Transgenicos, Quimicos, HigienismoDocument238 pagesComer y No Morir en El Intento-7 Ed - Mayo09-Aditivos, Cosmeticos, Transgenicos, Quimicos, Higienismocucocao100% (1)
- Informe de AnestesiaDocument12 pagesInforme de AnestesiaEdward AcunaNo ratings yet
- Instructivo de LimpiezaDocument14 pagesInstructivo de Limpiezajavier cordova flores100% (2)
- Wilhelm Reich Analisis Del Caracter PDFDocument403 pagesWilhelm Reich Analisis Del Caracter PDFShakur Shadyds100% (2)
- El Cuerpo Humano y Enfermedades Pantas Medicinales 63 PDFDocument111 pagesEl Cuerpo Humano y Enfermedades Pantas Medicinales 63 PDFIlianita MedinaNo ratings yet
- Linfaticos de Miembro Superior - CiucciDocument15 pagesLinfaticos de Miembro Superior - CiucciNicolas ValverdeNo ratings yet
- Appleton Accesorios 10Document2 pagesAppleton Accesorios 10Shodaime2No ratings yet
- Olga Cuevas Alimentos Que EquilibranDocument9 pagesOlga Cuevas Alimentos Que EquilibranDiana Árbol0% (1)
- GalvanismoDocument34 pagesGalvanismoEdgar Mauricio Arcos BeltránNo ratings yet
- Codo Muneca y ManoDocument42 pagesCodo Muneca y ManoTiitiimuaNo ratings yet
- LUMBDocument55 pagesLUMBAmos Coleman100% (1)
- Cardio 1Document9 pagesCardio 1Elizabeth RamírezNo ratings yet
- CIA General de AcerosDocument6 pagesCIA General de AcerosYesid Javier Martelo EllesNo ratings yet
- Kendall y KendallDocument3 pagesKendall y KendallSilvana D'Agosto100% (1)
- Onda corta diatermia aplicaciones tiposDocument1 pageOnda corta diatermia aplicaciones tiposMelisa Karen Garcia CayolaNo ratings yet
- Mapa Fisioterapia y Salud OcupacionalDocument1 pageMapa Fisioterapia y Salud OcupacionalJoselyn Danna100% (1)
- GALVANIZACIÓNDocument9 pagesGALVANIZACIÓNAlejandra AranaNo ratings yet
- Prueba de TrendelenburgDocument1 pagePrueba de TrendelenburgKendall Bartolo GarciaNo ratings yet
- Alto VoltajeDocument4 pagesAlto VoltajeKristina TabaresNo ratings yet
- Fundamentos Del ECGDocument12 pagesFundamentos Del ECGGbarielNo ratings yet
- Mapa Mental Del Tema Medula EspinalDocument1 pageMapa Mental Del Tema Medula EspinalMaría Fernanda EspinosaNo ratings yet
- Lesiones TendinosasDocument24 pagesLesiones TendinosasVanaii Ortiiz100% (1)
- Autonomic dysreflexia and hypertensive episode in spinal cord injury patientDocument4 pagesAutonomic dysreflexia and hypertensive episode in spinal cord injury patientJessica Sepúlveda SalasNo ratings yet
- Modelos del movimientoDocument16 pagesModelos del movimientoDanny PizarroNo ratings yet
- Corrientes ExitomotorasDocument16 pagesCorrientes ExitomotorasALISSON NAYELI MELGAR JAIMENo ratings yet
- Derrame ParaneumónicoDocument83 pagesDerrame ParaneumónicoMiguel Espinoza CarrilloNo ratings yet
- Pirámide Actividad FísicaDocument1 pagePirámide Actividad FísicaMaria Eugenia Valdes Planes100% (1)
- Sindrome de RaynaudDocument7 pagesSindrome de RaynaudDiana Lorena MEJIA MONJENo ratings yet
- CONTENIDO FISIOLOGIA 1 (Potencial de Acción Muscular)Document7 pagesCONTENIDO FISIOLOGIA 1 (Potencial de Acción Muscular)Nara FlosanNo ratings yet
- ElectroforesisDocument8 pagesElectroforesisSergio MéndezNo ratings yet
- Reflejos Primitivos Del Recien NacidoDocument1 pageReflejos Primitivos Del Recien Nacidoariel rios100% (2)
- Fibras Purkinje corazónDocument6 pagesFibras Purkinje corazónJeferson Marin TrejosNo ratings yet
- Ubicación Electrodos EKGDocument2 pagesUbicación Electrodos EKGFamiliaCubasAguirreNo ratings yet
- LUMBALGIASDocument44 pagesLUMBALGIASChristie ZamoraNo ratings yet
- Ii. Músculos de La Pared Abdominal ANTERIOR (Tabla 4-1) : AbdomenDocument60 pagesIi. Músculos de La Pared Abdominal ANTERIOR (Tabla 4-1) : AbdomengabrielaNo ratings yet
- Proyecto Osteoporosis 2018Document37 pagesProyecto Osteoporosis 2018Said GonzálezNo ratings yet
- Electrofisica de ElectroterapiaDocument6 pagesElectrofisica de ElectroterapiaNoé Jaime EstradaNo ratings yet
- Obstetricia-2020 UtpDocument1 pageObstetricia-2020 UtpLuciana Farfán MiñanNo ratings yet
- Analisis Caso Clinico EscoliosisDocument2 pagesAnalisis Caso Clinico EscoliosisFranco TabiloNo ratings yet
- Clase N 9 Anatomia de ToraxDocument28 pagesClase N 9 Anatomia de ToraxLuis Leon LizarbeNo ratings yet
- Ligadura de Trompas o TubariaDocument19 pagesLigadura de Trompas o TubariaFiorella RodriguezNo ratings yet
- Irrigación Sistema DigestivoDocument3 pagesIrrigación Sistema DigestivoBryan Castillo100% (1)
- Formato Diagnóstico Funcional de La CifDocument1 pageFormato Diagnóstico Funcional de La Cifalba0% (1)
- DeformacionDocument1 pageDeformaciononeydaNo ratings yet
- INTESTINO GRUESO QuirozDocument9 pagesINTESTINO GRUESO QuirozPamela Ramírez100% (1)
- Evolución UCEDocument11 pagesEvolución UCEPriscila NbNo ratings yet
- PALPACIÓN HIGADO BAZO PANCREASDocument35 pagesPALPACIÓN HIGADO BAZO PANCREASDouglas QuispeNo ratings yet
- Caso Clínico 04 - Sistema Nervioso PeriféricoDocument22 pagesCaso Clínico 04 - Sistema Nervioso PeriféricowillychirinosNo ratings yet
- Casos Clínicos Parte 4Document2 pagesCasos Clínicos Parte 4Juan José Gallo100% (1)
- Biomecánica de la Ventilación: Procesos, Fuerzas y Músculos InvolucradosDocument24 pagesBiomecánica de la Ventilación: Procesos, Fuerzas y Músculos InvolucradosAlvaro C Rivas VeraNo ratings yet
- Tercero Daysi Infografía TENS Ac2Document1 pageTercero Daysi Infografía TENS Ac2Alexandra TerceroNo ratings yet
- Sistemas de Captura y Análisis de Movimiento Cinemático Humano-Revisios SistematicaDocument11 pagesSistemas de Captura y Análisis de Movimiento Cinemático Humano-Revisios SistematicaTania MartínezNo ratings yet
- 09 Región CodoDocument38 pages09 Región CodomayhoyosNo ratings yet
- Actividad de Aplicación #04Document6 pagesActividad de Aplicación #04Mayra Alexandra Urquiaga MezaNo ratings yet
- Músculos de Cuello y TroncoDocument6 pagesMúsculos de Cuello y TroncoDominix SotoNo ratings yet
- Cinética y Cinemática Del TobilloDocument4 pagesCinética y Cinemática Del TobilloManuela PáezNo ratings yet
- AsmaDocument1 pageAsmaKarin Alvarez100% (1)
- 05 Corrientes RusasDocument2 pages05 Corrientes RusasCarlos GarciaNo ratings yet
- MedicinaDocument16 pagesMedicinaJuan Carlos CruzNo ratings yet
- Fracturas Tema de Exposicion Primeros Auxilios 2Document66 pagesFracturas Tema de Exposicion Primeros Auxilios 2Richard Bedón100% (1)
- Historia Clinica Psiquiatrica Mapa MentalDocument1 pageHistoria Clinica Psiquiatrica Mapa MentalEduardo CerquedaNo ratings yet
- Fractura de Maisonneuve: caso clínico de fractura espiral del peronéDocument19 pagesFractura de Maisonneuve: caso clínico de fractura espiral del peronéJesús Choque ZuritaNo ratings yet
- Enfermedades genéticas: causas, síntomas y tratamientosDocument3 pagesEnfermedades genéticas: causas, síntomas y tratamientosana cervantes0% (1)
- Via OlfatoriaDocument11 pagesVia OlfatoriaANA ESPINOSANo ratings yet
- Arritmias Fajuri 2009Document15 pagesArritmias Fajuri 2009jorge51988No ratings yet
- Electrofisiologia PDFDocument90 pagesElectrofisiologia PDFFabiola NorabuenaNo ratings yet
- La Sabiduria Del Agua de MarDocument64 pagesLa Sabiduria Del Agua de MarAna IsabelNo ratings yet
- Sanacion Con El Tarot - Ahimsa - 52 PáginasDocument371 pagesSanacion Con El Tarot - Ahimsa - 52 Páginas0vicman50% (8)
- PulmonDocument5 pagesPulmonDaniel Rojas GajardoNo ratings yet
- Agricultura Ecológica. El Huerto Medicinal. Pequeño Manual de Plantas Medicinales.Document58 pagesAgricultura Ecológica. El Huerto Medicinal. Pequeño Manual de Plantas Medicinales.Xose LopezNo ratings yet
- El Poder Nutritivo y Terapéutico Del AM 150710Document49 pagesEl Poder Nutritivo y Terapéutico Del AM 150710Jose PilottoNo ratings yet
- Valor Del Conocimiento en Cuanto A La AlimentacionDocument30 pagesValor Del Conocimiento en Cuanto A La Alimentacionrosica1973No ratings yet
- 84 Recetas Saludables para Quemar Grasa - Mariano OrzolaDocument237 pages84 Recetas Saludables para Quemar Grasa - Mariano Orzolalafercu100% (1)
- La Pesadilla de Los TransgénicosDocument8 pagesLa Pesadilla de Los Transgénicoseugenio_marti265No ratings yet
- Agua de Mar en La NutriciónDocument8 pagesAgua de Mar en La NutriciónDiana ÁrbolNo ratings yet
- Microbiota IntestinalDocument4 pagesMicrobiota Intestinaljesusolmo@hotmail.comNo ratings yet
- CFBRDocument30 pagesCFBRCarlos Fabian BurgueñoNo ratings yet
- 7 Probioticos y PrebioticosDocument17 pages7 Probioticos y PrebioticosSamuel de JesusNo ratings yet
- Agua de Mar Un Plasma Marino Al Alcance de TodosDocument57 pagesAgua de Mar Un Plasma Marino Al Alcance de Todosdianaprieto69100% (1)
- Tesina Xavier Coll ViolaDocument119 pagesTesina Xavier Coll Violadaisy_chavez_17No ratings yet
- Alteraciones Hormonales Asociadas A La NutriciónDocument60 pagesAlteraciones Hormonales Asociadas A La NutriciónEntrenamiento De Alto RendimientoNo ratings yet
- FenolftaleínaDocument2 pagesFenolftaleínaPerla HernandezNo ratings yet
- Tipos de AceroDocument2 pagesTipos de AceroJesús Hernández NavarroNo ratings yet
- Sesion 2 - Practica 2Document40 pagesSesion 2 - Practica 2Nicolás Campos PérezNo ratings yet
- Macromoléculas Introducción 2018-1Document44 pagesMacromoléculas Introducción 2018-1LAURA PINILLA AGATÓNNo ratings yet
- Producto Final Fiq-302Document18 pagesProducto Final Fiq-302ElisaNo ratings yet
- Informe RecristalizacionDocument5 pagesInforme RecristalizaciongoknaruNo ratings yet
- Ficha Tecnica Ac 4701-M-V2Document1 pageFicha Tecnica Ac 4701-M-V2Elmer Bueno VasquezNo ratings yet
- Formulación inorgánica: comparativa tradicional vs sistemática IUPACDocument1 pageFormulación inorgánica: comparativa tradicional vs sistemática IUPACEdward CullenNo ratings yet
- Materiales de ImpresiónDocument12 pagesMateriales de ImpresiónCeci SchulzNo ratings yet
- S2 - Hidrocarburos Alcanos y AlquenosDocument59 pagesS2 - Hidrocarburos Alcanos y AlquenosLourdes García GiraoNo ratings yet
- Interpretación y manejo de trastornos ácido-base: generalidades y primeras concepcionesDocument29 pagesInterpretación y manejo de trastornos ácido-base: generalidades y primeras concepcionesOscar Concha Jarpa0% (1)
- Síntesis Del Complejo Ioduro de (Diacuo) Bis (Etilendiamino) Cobre (II) (Cu (En) 2 (H2O) 2) I2.Document6 pagesSíntesis Del Complejo Ioduro de (Diacuo) Bis (Etilendiamino) Cobre (II) (Cu (En) 2 (H2O) 2) I2.nathaly310190No ratings yet
- Vida en Anaquel de Leche Chocolatada PDFDocument29 pagesVida en Anaquel de Leche Chocolatada PDFEdwin Chila ChoqueNo ratings yet
- Pavimento de Macadam AsfálticoDocument25 pagesPavimento de Macadam Asfálticoadelfofas60% (5)
- José Antonio Castro Cebrián - El Cementerio de La AlegríaDocument230 pagesJosé Antonio Castro Cebrián - El Cementerio de La AlegríaJaime Paul Bernal TolozaNo ratings yet
- Reacción ÁcidoDocument88 pagesReacción ÁcidoEdwin Nina MayhuaNo ratings yet
- Informe de ConstruccionDocument30 pagesInforme de ConstruccionjoseNo ratings yet
- Consulta 2Document4 pagesConsulta 2Monica MenaNo ratings yet
- S18 - Curso Refinación FCKDocument31 pagesS18 - Curso Refinación FCKRichard TineoNo ratings yet
- Zeina y Filmes Comestibles 2018Document3 pagesZeina y Filmes Comestibles 2018Daniel VélezNo ratings yet
- Carbonato de CobreDocument2 pagesCarbonato de Cobreandoba2013No ratings yet
- Unidad 2 Fuentes de LípidosDocument8 pagesUnidad 2 Fuentes de LípidoscarmenNo ratings yet
- IT-QAQC-ALSUD-04 Instructivo de Trabajo de Soldadura en TallerDocument7 pagesIT-QAQC-ALSUD-04 Instructivo de Trabajo de Soldadura en TallerAngelTinocoNo ratings yet
- Carta y Aplicaciones (Termodinámica)Document10 pagesCarta y Aplicaciones (Termodinámica)Mont's RmsNo ratings yet
- Clase 1 Introduccion NuevaDocument38 pagesClase 1 Introduccion Nuevajoseandres floresNo ratings yet
- INFORME de Supervision de ObrasDocument22 pagesINFORME de Supervision de ObrasKeswin Fabian Carrasco100% (1)
- MSDS Limpiador Desencrustante Serpentin - 36596900016 - BRILLALUM Ver2.5Document4 pagesMSDS Limpiador Desencrustante Serpentin - 36596900016 - BRILLALUM Ver2.5Marlon Pajaro GuzmanNo ratings yet