You might also like
- La Amenaza Del Caballo Oscuro - Marcus Sedgwick PDFDocument472 pagesLa Amenaza Del Caballo Oscuro - Marcus Sedgwick PDFMichael Danilo JimenezNo ratings yet
- Sal 7845Document128 pagesSal 7845Jesús Adrián Gómez OrtizNo ratings yet
- Antonio Velazco Pina La Mujer Dormida Debe Dar A LuzDocument119 pagesAntonio Velazco Pina La Mujer Dormida Debe Dar A LuzDiego Hoshin100% (5)
- La Cueva de Los Ocultos - Eva MorenoDocument61 pagesLa Cueva de Los Ocultos - Eva MorenoJesús Adrián Gómez OrtizNo ratings yet
- Declaracion FCDocument2 pagesDeclaracion FCJesús Adrián Gómez OrtizNo ratings yet
- Libro Blanco Europea 12Document18 pagesLibro Blanco Europea 12Jesús Adrián Gómez OrtizNo ratings yet
- Conjunto VocalDocument39 pagesConjunto Vocalsmg4704No ratings yet
- Principio de ParcimoniaDocument3 pagesPrincipio de ParcimoniaRaul Ernesto Lopez LopezNo ratings yet
- Thompson (Primer Grado)Document42 pagesThompson (Primer Grado)Jesús Adrián Gómez Ortiz100% (7)
- La Farmacia Comunitaria: Atención en Salud y Pluralismo AsistencialDocument13 pagesLa Farmacia Comunitaria: Atención en Salud y Pluralismo AsistencialJesús Adrián Gómez OrtizNo ratings yet
- Rosana Guber La Etnografia Metodo Campo Reflexividad Edicion 2011 PDFDocument151 pagesRosana Guber La Etnografia Metodo Campo Reflexividad Edicion 2011 PDFMario100% (2)
- BBPP 17 Colaboracion Farmacia Comunitaria DocenciaDocument12 pagesBBPP 17 Colaboracion Farmacia Comunitaria DocenciaJesús Adrián Gómez OrtizNo ratings yet
- 15 Nuevos Cantos para La Misa Carmelo ErdozainDocument31 pages15 Nuevos Cantos para La Misa Carmelo ErdozainJorge Narvaes86% (36)
- Alucinógenos y Cultura - Peter FurstDocument202 pagesAlucinógenos y Cultura - Peter Furstkatun11100% (15)
- I, Agem Profesional de Farmacias ComunitariasDocument14 pagesI, Agem Profesional de Farmacias ComunitariasJesús Adrián Gómez OrtizNo ratings yet
- Politica FarmaceuticaDocument164 pagesPolitica FarmaceuticaMalcomWallaceNo ratings yet
- Manual Operativo para El Manejo de Centros de AcopioDocument34 pagesManual Operativo para El Manejo de Centros de AcopioCarlos ChiribogaNo ratings yet
- Estudio PDFDocument8 pagesEstudio PDFJesús Adrián Gómez OrtizNo ratings yet
- Manual de Piano y Armonia Basica - CompletoDocument34 pagesManual de Piano y Armonia Basica - Completoapi-369880396% (136)
- Legislacion FarmaceuticaDocument10 pagesLegislacion FarmaceuticaJesús Adrián Gómez OrtizNo ratings yet
- Importante Farmacia Comunitaria y La Salud PublicaDocument6 pagesImportante Farmacia Comunitaria y La Salud Publicagbra80No ratings yet
- Farmacia ComunitariaDocument2 pagesFarmacia ComunitariaMaria Laura IamarinoNo ratings yet
- Importante Farmacia Comunitaria y La Salud PublicaDocument6 pagesImportante Farmacia Comunitaria y La Salud Publicagbra80No ratings yet
- Centro de Información de MedicamentosDocument14 pagesCentro de Información de MedicamentosJesús Adrián Gómez OrtizNo ratings yet
- 46 Educación SanitariaDocument59 pages46 Educación SanitariaEusebio Tocas MenaNo ratings yet
- Funciones Del FarmacéuticoDocument15 pagesFunciones Del FarmacéuticoJesús Adrián Gómez OrtizNo ratings yet
- Tamiz Neonatal AmpliadoDocument2 pagesTamiz Neonatal AmpliadoJesús Adrián Gómez OrtizNo ratings yet
- Tercer Consenso de Granada Sobre La Atencion FarmaceuticaDocument13 pagesTercer Consenso de Granada Sobre La Atencion Farmaceuticasandra milenaNo ratings yet
- Depresion en El Embarazo y PospartoDocument8 pagesDepresion en El Embarazo y PospartoJesús Adrián Gómez OrtizNo ratings yet
- Sonidos Del SilencioDocument3 pagesSonidos Del SilencioJesús Adrián Gómez OrtizNo ratings yet
- Analisis Del Proseso ConstructivoDocument9 pagesAnalisis Del Proseso ConstructivoJose de RosasNo ratings yet
- Reporte de Dengue en Ucayali 2013Document15 pagesReporte de Dengue en Ucayali 2013FerdieNo ratings yet
- Matemáticas IDocument285 pagesMatemáticas Ielsupernino50% (6)
- Inmunodeficiencia Variable ComúnDocument1 pageInmunodeficiencia Variable ComúnPaola RochaNo ratings yet
- Taller4 2023 1Document5 pagesTaller4 2023 1alex quinteroNo ratings yet
- Psicología Del Aprendizaje - Sem 2Document54 pagesPsicología Del Aprendizaje - Sem 2Sueños de LunaNo ratings yet
- Pearson Resolucion Del Capitulo 8Document10 pagesPearson Resolucion Del Capitulo 8Gian Piero Marcalaya FernándezNo ratings yet
- Caso Estudio - Arquitectura de SoftwareDocument8 pagesCaso Estudio - Arquitectura de SoftwareJulio Alberto Reinoso VillarroelNo ratings yet
- Manual Autoridad Certificado de PromociónDocument11 pagesManual Autoridad Certificado de PromociónSammy AlquingaNo ratings yet
- Bianco 2 ESPAÑOL 6 GRADODocument19 pagesBianco 2 ESPAÑOL 6 GRADOBolivar Concepcion PrescottNo ratings yet
- Com Tarea1Document21 pagesCom Tarea1mac_ejbs100% (1)
- Ciclo Rankine Ejercicios ResueltosDocument7 pagesCiclo Rankine Ejercicios ResueltosIsa CardonaNo ratings yet
- Cualidades Estèticas de La QuinchaDocument6 pagesCualidades Estèticas de La QuinchaNicoll RodriguezNo ratings yet
- El KerógenoDocument20 pagesEl KerógenoLuis Alberto Tupa CoilaNo ratings yet
- Seminario Anomalias Del Aparato Genital Femenino Katherine Leiva SalazarDocument39 pagesSeminario Anomalias Del Aparato Genital Femenino Katherine Leiva SalazarIrina Surco RodríguezNo ratings yet
- Evaluacion y Acreditacion de La Educación Superior en MexicoDocument23 pagesEvaluacion y Acreditacion de La Educación Superior en MexicoCabrera YesLopNo ratings yet
- Ángulos ++Document16 pagesÁngulos ++Jesús GarcíaNo ratings yet
- Ayudantía N°3.1 - Ing PDFDocument1 pageAyudantía N°3.1 - Ing PDFsdfghfgmjhfgdfdsNo ratings yet
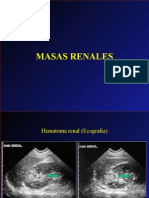
- Masas - Renales - Imagen DiagnósticaDocument22 pagesMasas - Renales - Imagen DiagnósticaramonNo ratings yet
- Codigo Tributario Libro IDocument18 pagesCodigo Tributario Libro IRichard Ramos CutimboNo ratings yet
- Municipio de ChimalhuacánDocument94 pagesMunicipio de ChimalhuacánMatias Cruz HumbertoNo ratings yet
- Indicadores en Música ProfesoresDocument45 pagesIndicadores en Música ProfesoresfelicreazNo ratings yet
- La Omc y La IccDocument5 pagesLa Omc y La IccLeomarys PaulinoNo ratings yet
- Diagrama de Flujo de Proceso de Produccion de Un LapizDocument1 pageDiagrama de Flujo de Proceso de Produccion de Un LapizadrianNo ratings yet
- Simulacion de Procesos Quimicos: Universidad Nacional Del Altiplano Facultad de Ingenieria QuimicaDocument19 pagesSimulacion de Procesos Quimicos: Universidad Nacional Del Altiplano Facultad de Ingenieria Quimicaelia melayde rosello ccasaNo ratings yet
- Trabajo Final - Crucigrama Con RespuestasDocument2 pagesTrabajo Final - Crucigrama Con RespuestasTami LiendoNo ratings yet
- Isomeros y EstereoquimicaDocument3 pagesIsomeros y EstereoquimicaManuel Alejandro PeñafielNo ratings yet
- S8 S2-MonopolioDocument23 pagesS8 S2-MonopolioSalem IsraNo ratings yet
- Definición de Eficiencia: Optimizar Recursos para Lograr MetasDocument3 pagesDefinición de Eficiencia: Optimizar Recursos para Lograr MetasRosa Luz Chavez Lopez86% (7)
- Lista de Alarmas CarrierDocument16 pagesLista de Alarmas CarrierFABIAN ZARATE RAMOS100% (2)