You might also like
- The Slim Book of Health Pearls: Hormones, Nerves, and StressFrom EverandThe Slim Book of Health Pearls: Hormones, Nerves, and StressNo ratings yet
- Dermatological and Haemato-Biochemical Characteristics of Canine HypothyroidismDocument5 pagesDermatological and Haemato-Biochemical Characteristics of Canine HypothyroidismsumNo ratings yet
- CANINE-Pathophysiology of Organ Failure in Severe Acute Pancreatitis in DogsDocument10 pagesCANINE-Pathophysiology of Organ Failure in Severe Acute Pancreatitis in Dogstaner_soysurenNo ratings yet
- Pain Book FeinDocument102 pagesPain Book FeinHansen NielsNo ratings yet
- VPK3611 Diseases of The Nervous System Seizures: Intan ShafieDocument29 pagesVPK3611 Diseases of The Nervous System Seizures: Intan Shafieiloveyou_143No ratings yet
- Skinner's Behavioral Approach to PersonalityDocument6 pagesSkinner's Behavioral Approach to PersonalityRansuu GamesNo ratings yet
- Fluoxetine Combined With Clorazepate Dipotassium and BehaviourDocument5 pagesFluoxetine Combined With Clorazepate Dipotassium and BehaviourAngélica Lozano CastroNo ratings yet
- Veterinary Neurology Symposium: Approach to Seizure PatientsDocument9 pagesVeterinary Neurology Symposium: Approach to Seizure PatientsOzkar JaraNo ratings yet
- Atlas of Small Animal Wound Management and Reconstructive SurgeryFrom EverandAtlas of Small Animal Wound Management and Reconstructive SurgeryNo ratings yet
- Urinary Catheter Placement For Feline ObstructionDocument6 pagesUrinary Catheter Placement For Feline ObstructionWilliam ChandlerNo ratings yet
- Canine InsulinomaDocument5 pagesCanine Insulinomasoff4ikaNo ratings yet
- Neurological ExamDocument6 pagesNeurological ExamTiti Daraban100% (2)
- Animal Training and ImplicationsDocument12 pagesAnimal Training and ImplicationsBarbaraNo ratings yet
- What Is Functional NeurologyDocument2 pagesWhat Is Functional NeurologydrtimadamsNo ratings yet
- Hip Dysplasia in Dogs: A Guide For Dog OwnersDocument6 pagesHip Dysplasia in Dogs: A Guide For Dog Ownerstaner_soysurenNo ratings yet
- Vet PathologyDocument128 pagesVet PathologyMegarsaNo ratings yet
- Olfaction PDFDocument5 pagesOlfaction PDFjorgegrodl100% (1)
- Diagnostic Touch Its Principles and Application PDFDocument9 pagesDiagnostic Touch Its Principles and Application PDFJakobStierleNo ratings yet
- Biological RhythmsDocument4 pagesBiological RhythmsMarco Alfieri100% (1)
- Dermatology For The Small Animal Practitioner (Made Easy Series)Document163 pagesDermatology For The Small Animal Practitioner (Made Easy Series)Nela Frade0% (1)
- Practical Transfusion Medicine for the Small Animal PractitionerFrom EverandPractical Transfusion Medicine for the Small Animal PractitionerNo ratings yet
- Enzymes by CatheyDocument27 pagesEnzymes by CatheyChad Ashley VandenbergNo ratings yet
- Physical Examination of The Lymphatic System: Russell C Hendershot DO, MS, FAAFPDocument50 pagesPhysical Examination of The Lymphatic System: Russell C Hendershot DO, MS, FAAFPNajwa100% (1)
- CCJM Essential Tremor Choosing The Right Management Plan For Your PatientDocument8 pagesCCJM Essential Tremor Choosing The Right Management Plan For Your PatientBrian HarrisNo ratings yet
- Mucocele 2Document13 pagesMucocele 2FEY YENG SOFIA FUNG GUADALUPENo ratings yet
- IBRO-APRC Global Advocacy 2015 Powerpoint PresentationsDocument328 pagesIBRO-APRC Global Advocacy 2015 Powerpoint PresentationsInternational Brain Research OrganizationNo ratings yet
- Suture MaterialDocument7 pagesSuture Materialtaner_soysuren100% (1)
- Duncan and Prasse's Veterinary Laboratory Medicine: Clinical PathologyFrom EverandDuncan and Prasse's Veterinary Laboratory Medicine: Clinical PathologyKenneth S. LatimerNo ratings yet
- Carotid Artery Dissection PDFDocument3 pagesCarotid Artery Dissection PDFOdranoelRichamNo ratings yet
- Heartworm Disease in DogsDocument5 pagesHeartworm Disease in DogsDumapis RichardNo ratings yet
- Horner's Syndrome: Lid) Eye) (" ")Document3 pagesHorner's Syndrome: Lid) Eye) (" ")D TNo ratings yet
- Neural Mechanisms of Salivary Gland SecretionDocument247 pagesNeural Mechanisms of Salivary Gland SecretionMassimo Baudo100% (1)
- NeurolepticsDocument6 pagesNeurolepticsVantaku Krishna Swamy NaiduNo ratings yet
- Hills Atlas of Clinical AnatomyDocument104 pagesHills Atlas of Clinical AnatomykritvetNo ratings yet
- Current Diagnostic Techniques in Veterinary Surgery PDFDocument2 pagesCurrent Diagnostic Techniques in Veterinary Surgery PDFKirti JamwalNo ratings yet
- Brainstem Control of Spinal Cord FunctionFrom EverandBrainstem Control of Spinal Cord FunctionCharles D. BarnesRating: 3.5 out of 5 stars3.5/5 (14)
- Pseudo Pregnancy in DogsDocument11 pagesPseudo Pregnancy in DogsKay ChuaNo ratings yet
- Xylazine Sedation Antagonized With TolazolineDocument9 pagesXylazine Sedation Antagonized With Tolazolinetaner_soysurenNo ratings yet
- Veterinary Critical CareDocument15 pagesVeterinary Critical Caresudanfx100% (1)
- 2011 Reproductive Cycles of The Domestic BitchDocument11 pages2011 Reproductive Cycles of The Domestic BitchMariaCamilaLeonPerezNo ratings yet
- VEM5384 Clincial Neurology DisordersDocument62 pagesVEM5384 Clincial Neurology DisordersdeadnarwhalNo ratings yet
- Veterinary Drug IndexDocument335 pagesVeterinary Drug IndexJulienne Frances De CastroNo ratings yet
- Reflex ArcDocument16 pagesReflex ArcNiaz Ahammed50% (2)
- Fluorescent Light Full-SpectrumDocument15 pagesFluorescent Light Full-SpectrumAlexander WunschNo ratings yet
- 10th World Veterinary Dental Congress 2007 BrazilDocument127 pages10th World Veterinary Dental Congress 2007 BrazilHumberto FustesNo ratings yet
- Small Animal Spinal Disorders Diagnosis and Surgery, 2nd Edition (Vetbooks - Ir)Document369 pagesSmall Animal Spinal Disorders Diagnosis and Surgery, 2nd Edition (Vetbooks - Ir)ghimboasaNo ratings yet
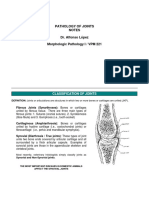
- Pathology of JointsDocument13 pagesPathology of JointswobblegobbleNo ratings yet
- Status Epilepticus-Patient Management and Pharmocologic TheraphyDocument7 pagesStatus Epilepticus-Patient Management and Pharmocologic Theraphytaner_soysuren100% (1)
- Canine Separation Anxiety - Strategies For Treatment and ManagementDocument9 pagesCanine Separation Anxiety - Strategies For Treatment and ManagementAnonymous TDI8qdYNo ratings yet
- Top 5 Leukogram PatternsDocument3 pagesTop 5 Leukogram PatternsSandraHermosilloNo ratings yet
- Veterinary Acupuncturejshqkjq V1aDocument2 pagesVeterinary Acupuncturejshqkjq V1aGheorgian HageNo ratings yet
- FELINE-Clinical Forms of Acquired Myasthenia Gravis in CatsDocument7 pagesFELINE-Clinical Forms of Acquired Myasthenia Gravis in Catstaner_soysurenNo ratings yet
- Endocrine Glands PDFDocument99 pagesEndocrine Glands PDFXochitl ZambranoNo ratings yet
- Applied Animal EndocrinologyDocument252 pagesApplied Animal Endocrinologylulignani100% (1)
- The Pattern Approach To Logic DiagnosisDocument12 pagesThe Pattern Approach To Logic Diagnosistaner_soysuren100% (1)
- Stages of Sleep, Dreams, and Sleep ApneaDocument16 pagesStages of Sleep, Dreams, and Sleep ApneaMarina Los100% (1)
- JULAUG 2014 Selected PagesDocument5 pagesJULAUG 2014 Selected PagesStephen PopeNo ratings yet
- Vet CareplanexampleDocument6 pagesVet CareplanexampleAnonymous eJZ5HcNo ratings yet
- Two-Dimensional Gel Electrophoresis of Proteins: Methods and ApplicationsFrom EverandTwo-Dimensional Gel Electrophoresis of Proteins: Methods and ApplicationsJulio CelisNo ratings yet
- Xylazine Sedation Antagonized With TolazolineDocument9 pagesXylazine Sedation Antagonized With Tolazolinetaner_soysurenNo ratings yet
- Web MedDocument3 pagesWeb Medtaner_soysurenNo ratings yet
- Veterinary Medical Resources On The INternetDocument7 pagesVeterinary Medical Resources On The INternettaner_soysuren100% (1)
- Ultrasonography of The EyeDocument8 pagesUltrasonography of The Eyetaner_soysurenNo ratings yet
- Small Animal/ExoticsDocument4 pagesSmall Animal/Exoticstaner_soysuren100% (1)
- Tracheostomy Techniques and ManagementDocument9 pagesTracheostomy Techniques and Managementtaner_soysuren100% (1)
- Three-Dimensional Computed Tomography-User - Friendly ImagesDocument5 pagesThree-Dimensional Computed Tomography-User - Friendly Imagestaner_soysurenNo ratings yet
- Veterinary OncologyDocument3 pagesVeterinary Oncologytaner_soysuren100% (1)
- Veterinary ControversiesDocument3 pagesVeterinary Controversiestaner_soysurenNo ratings yet
- Treating Bone Deformities With Circular External Skeletal FixationDocument10 pagesTreating Bone Deformities With Circular External Skeletal Fixationtaner_soysurenNo ratings yet
- Small Animal/ExoticsDocument5 pagesSmall Animal/Exoticstaner_soysurenNo ratings yet
- Systemic Absorption of Topically Administered DrugsDocument8 pagesSystemic Absorption of Topically Administered Drugstaner_soysuren100% (2)
- Traditional Anti Fungal Logic AgentsDocument6 pagesTraditional Anti Fungal Logic Agentstaner_soysuren100% (1)
- The Expanding Role of Veterinary TechniciansDocument3 pagesThe Expanding Role of Veterinary Technicianstaner_soysurenNo ratings yet
- The Pattern Approach To Logic DiagnosisDocument12 pagesThe Pattern Approach To Logic Diagnosistaner_soysuren100% (1)
- Susceptibility of Fleas To Control AgentsDocument2 pagesSusceptibility of Fleas To Control Agentstaner_soysurenNo ratings yet
- Supracondylar Femoral Fractures in Adult AnimalsDocument12 pagesSupracondylar Femoral Fractures in Adult Animalstaner_soysuren100% (3)
- Suture MaterialDocument7 pagesSuture Materialtaner_soysuren100% (1)
- Surgical Techniques For Extra Vascular Occlusiion of Tic ShuntsDocument7 pagesSurgical Techniques For Extra Vascular Occlusiion of Tic Shuntstaner_soysurenNo ratings yet
- Status Epilepticus-Clinical Features and Pa Tho PhysiologyDocument8 pagesStatus Epilepticus-Clinical Features and Pa Tho Physiologytaner_soysurenNo ratings yet
- Status Epilepticus-Managing Refractory Cases and Treating Out-Of-hospital PatientsDocument7 pagesStatus Epilepticus-Managing Refractory Cases and Treating Out-Of-hospital Patientstaner_soysurenNo ratings yet
- Status Epilepticus-Patient Management and Pharmocologic TheraphyDocument7 pagesStatus Epilepticus-Patient Management and Pharmocologic Theraphytaner_soysuren100% (1)
- Soft Tissue SurgeryDocument3 pagesSoft Tissue Surgerytaner_soysuren0% (1)
- SOW-Feeding Management During Sow LactationDocument6 pagesSOW-Feeding Management During Sow Lactationtaner_soysuren100% (1)
- SALMON FİSH-Performing Diagnostic Procedures On Salmonid FishesDocument7 pagesSALMON FİSH-Performing Diagnostic Procedures On Salmonid Fishestaner_soysurenNo ratings yet
- Small Animal/ExoticsDocument3 pagesSmall Animal/Exoticstaner_soysurenNo ratings yet
- Small Animal Oxygen TherapyDocument10 pagesSmall Animal Oxygen Therapytaner_soysuren100% (1)
- Should I Recommend The Use of Oral GlycosaminoglycansDocument2 pagesShould I Recommend The Use of Oral Glycosaminoglycanstaner_soysurenNo ratings yet
- Role of Glutamine in Health and DiseaseDocument8 pagesRole of Glutamine in Health and Diseasetaner_soysurenNo ratings yet
- The AUTO-bıography of Aziz SancarDocument25 pagesThe AUTO-bıography of Aziz SancarasliNo ratings yet
- Exam+1+BIOL202+SS2+14 +KEY-2Document8 pagesExam+1+BIOL202+SS2+14 +KEY-2Haley HamillNo ratings yet
- Cell CultureDocument12 pagesCell CultureNihmathullah Mohamed RizwanNo ratings yet
- Blood PhysiologyDocument17 pagesBlood Physiologymuhammad sadiqNo ratings yet
- Intro Microbiology Study Microscopic LifeDocument13 pagesIntro Microbiology Study Microscopic LifeVon Valentine MhuteNo ratings yet
- Causes of Migraine PDFDocument7 pagesCauses of Migraine PDFJohn Christopher LucesNo ratings yet
- ApoptosisDocument35 pagesApoptosisDR VIJAY MARAKALA50% (2)
- Roles of Vitamins in Stem Cells: Cellular and Molecular Life Sciences May 2020Document22 pagesRoles of Vitamins in Stem Cells: Cellular and Molecular Life Sciences May 2020Chanda DajaniNo ratings yet
- Epidemiology of Brain TumorsDocument25 pagesEpidemiology of Brain Tumorsinternship unsyiahNo ratings yet
- Theories of AgingDocument4 pagesTheories of AgingJozelle Marie MartinezNo ratings yet
- Stevens (Science) Med TechDocument31 pagesStevens (Science) Med TechnotsoninjaninjaNo ratings yet
- 05-02-Genetika MikroorganismeDocument48 pages05-02-Genetika MikroorganismeKinad DanikNo ratings yet
- Restitution: Over Correction Because The Corrective Action Goes Beyond The Disruption or Damage Caused byDocument5 pagesRestitution: Over Correction Because The Corrective Action Goes Beyond The Disruption or Damage Caused byiulia9gavrisNo ratings yet
- Dr. Budhi Setianto Dyslipidemia and AtheroclerosisDocument25 pagesDr. Budhi Setianto Dyslipidemia and AtheroclerosisYunita WidyaningsihNo ratings yet
- 4727 Whatman DNA - OmniSwab Appnote - v15Document6 pages4727 Whatman DNA - OmniSwab Appnote - v15Alejandra Torres100% (1)
- Volpe's Understanding EvolutionDocument303 pagesVolpe's Understanding EvolutionAnonymous kg7YBMFH100% (1)
- 3rd Quarter Exam Science 10Document3 pages3rd Quarter Exam Science 10Evelyn71% (7)
- Niper 2.Document42 pagesNiper 2.GANESH KUMAR JELLANo ratings yet
- مختبر مايكرو 2و 3 و 4 PDFDocument13 pagesمختبر مايكرو 2و 3 و 4 PDFAhmed Assaf100% (1)
- IEL / ETM Case Study Series: Xeno Mouse Patent Portfolio AnalysisDocument83 pagesIEL / ETM Case Study Series: Xeno Mouse Patent Portfolio AnalysisGunaseelan RamasamyNo ratings yet
- 5 Health As A Social InstitutionDocument34 pages5 Health As A Social InstitutionSze Yinn PikaaNo ratings yet
- Cell Communication PDFDocument109 pagesCell Communication PDFediaz_956003100% (1)
- Glomerular Diseases: Assistant Professor Dr. Shumaila Rafi MedicineDocument17 pagesGlomerular Diseases: Assistant Professor Dr. Shumaila Rafi MedicineMuhammad MakkiNo ratings yet
- Coagulation DisordersDocument26 pagesCoagulation DisordersLia pramita0% (1)
- Hair BiologyDocument16 pagesHair BiologyTakis VasilopoulosNo ratings yet
- Nephrology MnemonicsDocument5 pagesNephrology MnemonicsAnnapurna DangetiNo ratings yet
- PHQ - Patient Health Questionnaire: Add ColumnsDocument1 pagePHQ - Patient Health Questionnaire: Add ColumnsFar SultanNo ratings yet
- Cre-Lox recombination system for targeted DNA modificationDocument6 pagesCre-Lox recombination system for targeted DNA modificationanirbanmanna88320No ratings yet
- Fluid & Electrolytes ManagementDocument18 pagesFluid & Electrolytes ManagementSh ReyaNo ratings yet
- Developmental Biology BY1101Document8 pagesDevelopmental Biology BY1101Dr. Kaushal Kishor SharmaNo ratings yet