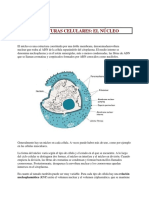
You might also like
- Tema. Bioquimica Acidos NucleicosDocument9 pagesTema. Bioquimica Acidos Nucleicosmgonate100% (1)
- Nucleótidos y AdnDocument14 pagesNucleótidos y AdnRichivillNo ratings yet
- Ácidos Nucleicos Biología I 3 Clase 27 OctDocument39 pagesÁcidos Nucleicos Biología I 3 Clase 27 Octsora polar bearNo ratings yet
- Tema CincoDocument5 pagesTema Cincomaria barrera juradoNo ratings yet
- Ácidos NucleicosDocument6 pagesÁcidos NucleicosMaríaNo ratings yet
- Ensayo 7. NucleotidosDocument10 pagesEnsayo 7. NucleotidosManuel AntonioNo ratings yet
- ÁCIDOS NUCLEICOS Apuntes PDFDocument10 pagesÁCIDOS NUCLEICOS Apuntes PDFProf. Víctor M. VitoriaNo ratings yet
- T5 Los Ácidos NucleicosDocument4 pagesT5 Los Ácidos Nucleicosaitana.roso06No ratings yet
- Acidos NucleicosDocument6 pagesAcidos Nucleicospuchiti1234No ratings yet
- Resumen Final Biol. Cel. y Molec.Document211 pagesResumen Final Biol. Cel. y Molec.Hugo MedusNo ratings yet
- Cuestionario 4 de Ácidos Nucléicos GLEICE KEILA 25042022Document4 pagesCuestionario 4 de Ácidos Nucléicos GLEICE KEILA 25042022amissadayNo ratings yet
- Bases Moleculares de La Información GenéticaDocument13 pagesBases Moleculares de La Información GenéticaIrene Sánchez LouNo ratings yet
- Taller Biología MolecularDocument9 pagesTaller Biología MolecularJose E. MirandaNo ratings yet
- Ácidos NucléicosDocument11 pagesÁcidos NucléicosJavier SosaNo ratings yet
- Tema 6 Ácidos NucleicosDocument11 pagesTema 6 Ácidos NucleicosPepe Gomez bernalNo ratings yet
- Acidos NucleicosDocument20 pagesAcidos NucleicosMarianela Rios AillonNo ratings yet
- Estructura de Los Ácidos NucleicosDocument5 pagesEstructura de Los Ácidos NucleicosYesid HernandezNo ratings yet
- T 6 Ac Nucleicos 1 11Document11 pagesT 6 Ac Nucleicos 1 11marito21777No ratings yet
- Tema 7. Acidos NucleicosDocument37 pagesTema 7. Acidos NucleicosLaorden LucíaNo ratings yet
- Acidos NucleicosDocument21 pagesAcidos NucleicosFranco Apaza AlaveNo ratings yet
- Nucleósidos 2Document10 pagesNucleósidos 2Arian CantillanoNo ratings yet
- Acidos NucleicosDocument3 pagesAcidos Nucleicosgarnicadaniela38No ratings yet
- T 6 Ac NucleicosDocument12 pagesT 6 Ac NucleicosAlejandro AlemanNo ratings yet
- 2023 Bioquímica 4 Ácidos Nucleicos - Clasificación y Estructura.Document34 pages2023 Bioquímica 4 Ácidos Nucleicos - Clasificación y Estructura.Fabiana Valentina Contreras ElguetaNo ratings yet
- Lucha contra corrupción e impunidadDocument14 pagesLucha contra corrupción e impunidadPoveda Multiservicios100% (1)
- Acidos NucleicosDocument7 pagesAcidos NucleicosamandaNo ratings yet
- Tema 1 Genetica Molecular DEFDocument10 pagesTema 1 Genetica Molecular DEFAbraham alcalaNo ratings yet
- 4 Acidos NucleicosDocument8 pages4 Acidos Nucleicoscyber pgNo ratings yet
- Biosintesis AcidosDocument15 pagesBiosintesis AcidosLuis Fernando Porras ZapataNo ratings yet
- Dna y RnaDocument5 pagesDna y RnaVictor PintleNo ratings yet
- NUCLEÓTIDOSDocument5 pagesNUCLEÓTIDOSGaby_biologia83% (6)
- Tema 6 Acidos NucleicosDocument12 pagesTema 6 Acidos NucleicosDavid MorenoNo ratings yet
- Anexo 3Document24 pagesAnexo 3MARIA LUISA CHACCA BUTRONNo ratings yet
- Separata - Acidos NucleicosDocument8 pagesSeparata - Acidos NucleicosCésar AlejandroNo ratings yet
- ADN Estructura y FunciónDocument11 pagesADN Estructura y FunciónPabloNo ratings yet
- Acidos NucleicosDocument52 pagesAcidos Nucleicosquimicoupagu96% (23)
- Los Nucleótidos y Los Ácidos NucléicosDocument7 pagesLos Nucleótidos y Los Ácidos NucléicoswiritaNo ratings yet
- Tema 4 - Biología MolecularDocument3 pagesTema 4 - Biología MolecularMari Carmen ReinéNo ratings yet
- Ácidos NucleicosDocument8 pagesÁcidos NucleicosRaquel García HortalNo ratings yet
- 2 - Estructura Adn y ArnDocument17 pages2 - Estructura Adn y ArnEl Profe de BiologíaNo ratings yet
- Tema 25Document8 pagesTema 25Elena ManchonNo ratings yet
- Ácidos NucleicosDocument25 pagesÁcidos NucleicosLola100% (1)
- Estructura y composición de los ácidos nucleicos (ADN y ARNDocument23 pagesEstructura y composición de los ácidos nucleicos (ADN y ARNDalia MonteagudoNo ratings yet
- Quím de Ácidos NucleicosDocument10 pagesQuím de Ácidos NucleicosLizbeth Oyarce OrtizNo ratings yet
- Biología MolecularDocument30 pagesBiología MolecularAlejandra GonzalezNo ratings yet
- Biología Tema 5 Ácidos NucleicosDocument3 pagesBiología Tema 5 Ácidos NucleicosAnaNo ratings yet
- Ácidos NucleicosDocument41 pagesÁcidos NucleicosMary UmaquingaNo ratings yet
- TEMA 3 Ácidos NucleicosDocument7 pagesTEMA 3 Ácidos NucleicosElena Sabas CarvajalNo ratings yet
- GUIA SOBRE ESTRUCTURA Y FUNCION DEL ADNDocument20 pagesGUIA SOBRE ESTRUCTURA Y FUNCION DEL ADNMaryLaderaMagoNo ratings yet
- Estructura del DNADocument22 pagesEstructura del DNAJonatan DuranNo ratings yet
- Ácidos NucleicosDocument10 pagesÁcidos NucleicosJavier Marquez HurtadoNo ratings yet
- Nucleótidos y Ácidos NucleicosDocument9 pagesNucleótidos y Ácidos NucleicosLourdes SoriaNo ratings yet
- ACIDOS NUCLEICOS. OcthDocument10 pagesACIDOS NUCLEICOS. OcthWilmerNo ratings yet
- Biología Celular y Molecular 1Document75 pagesBiología Celular y Molecular 1Christian AvendañoNo ratings yet
- Biologia AgusDocument10 pagesBiologia AgusRodrigo LuceroNo ratings yet
- Guía 2 ADN y ARN (4° Medio)Document6 pagesGuía 2 ADN y ARN (4° Medio)Andres Oyanadel CortesNo ratings yet
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012From EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012No ratings yet
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSFrom EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSRating: 5 out of 5 stars5/5 (1)
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017From EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017No ratings yet
- Caratula San PedroDocument2 pagesCaratula San PedroIris LissiNo ratings yet
- Mitos BarrancaDocument3 pagesMitos BarrancaIris LissiNo ratings yet
- Noticia de BarrancaDocument2 pagesNoticia de BarrancaIris LissiNo ratings yet
- Valor Temporal Del DineroDocument1 pageValor Temporal Del DineroIris LissiNo ratings yet
- Carta de RenunciaDocument2 pagesCarta de RenunciaIris LissiNo ratings yet
- Faisan de OroDocument1 pageFaisan de OroIris LissiNo ratings yet
- Mapa BarrancaDocument4 pagesMapa BarrancaIris LissiNo ratings yet
- PROFORMADocument1 pagePROFORMAIris LissiNo ratings yet
- Región CostaDocument1 pageRegión CostaIris LissiNo ratings yet
- Letras - Periodico MuralDocument6 pagesLetras - Periodico MuralIris Lissi0% (1)
- Empresa de Transporte y ServiciosDocument1 pageEmpresa de Transporte y ServiciosIris LissiNo ratings yet
- CurriculumDocument6 pagesCurriculumIris LissiNo ratings yet
- FotosheckDocument2 pagesFotosheckIris LissiNo ratings yet
- MicroprocesadorDocument4 pagesMicroprocesadorIris LissiNo ratings yet
- AcrósticoDocument7 pagesAcrósticoIris LissiNo ratings yet
- Carta de GarantiaDocument2 pagesCarta de GarantiaIris LissiNo ratings yet
- PapaDocument1 pagePapaIris LissiNo ratings yet
- Propiedades de Los MaterialesDocument10 pagesPropiedades de Los MaterialesIris LissiNo ratings yet
- Contrato de CompraDocument2 pagesContrato de CompraIris Lissi100% (5)
- Letras para CuadernosDocument4 pagesLetras para CuadernosIris LissiNo ratings yet
- Acta de EntregaDocument1 pageActa de EntregaIris LissiNo ratings yet
- Formato de AsistenciaDocument1 pageFormato de AsistenciaIris LissiNo ratings yet
- Acta de CompromisoDocument2 pagesActa de CompromisoIris LissiNo ratings yet
- Acta de CompromisoDocument2 pagesActa de CompromisoIris LissiNo ratings yet
- Vacuna AntiedadDocument2 pagesVacuna AntiedadRicardo WeschenfellerNo ratings yet
- Biologia Psicologia y Sociologia Intromisiones y RDocument16 pagesBiologia Psicologia y Sociologia Intromisiones y RMargaret Abreu100% (1)
- BacteriasDocument16 pagesBacteriassergioNo ratings yet
- CarbohidratosDocument4 pagesCarbohidratosFabian lozano ballesterosNo ratings yet
- Reproducción asexual enDocument5 pagesReproducción asexual enGabrielNo ratings yet
- Clase 1 ZoologiaDocument10 pagesClase 1 ZoologiaSandra Milena Guerra HernándezNo ratings yet
- E ColiDocument8 pagesE ColiLuis HurtadoNo ratings yet
- Aplicación Del Método CientíficoDocument5 pagesAplicación Del Método Científicote adoro G.AvilaNo ratings yet
- O GlicosilaciónDocument4 pagesO GlicosilaciónKito TongHuiNo ratings yet
- Libro BioquimicaDocument270 pagesLibro BioquimicaBoris SalasNo ratings yet
- HONGOSDocument5 pagesHONGOSmarisol salgado perezNo ratings yet
- Enzima de Restricción, Adn Ligasa y Adn RecombinanteDocument15 pagesEnzima de Restricción, Adn Ligasa y Adn RecombinanteMARILYN ISABEL ORTIZ ESPIRITUNo ratings yet
- Tejido Conectivo LaxoDocument6 pagesTejido Conectivo LaxoRox SalazarNo ratings yet
- Desarrollo embrionario en la primera semanaDocument2 pagesDesarrollo embrionario en la primera semanaSebastian JimenezNo ratings yet
- El Reino PlantaeDocument2 pagesEl Reino PlantaeRosalía Huamán QuiñonezNo ratings yet
- Estructura nuclearDocument5 pagesEstructura nuclearAndres Felipe Prieto AlarconNo ratings yet
- Neuronas piramidales: funciones clave en el cerebroDocument4 pagesNeuronas piramidales: funciones clave en el cerebromayNo ratings yet
- Laboratorio Partes de La Flor Octavo.Document2 pagesLaboratorio Partes de La Flor Octavo.juan camilo ladino caceresNo ratings yet
- Sexualidad ResponsableDocument3 pagesSexualidad ResponsableErwin PaNo ratings yet
- Guía para Examen Ordinario de Ética Sociedad y TecnologíaDocument3 pagesGuía para Examen Ordinario de Ética Sociedad y TecnologíaEsthela FloresNo ratings yet
- Capitulo Muestra-Biologia BachilleratoDocument25 pagesCapitulo Muestra-Biologia Bachilleratojuan nanesNo ratings yet
- Tarea de Biologia de Ada 1Document11 pagesTarea de Biologia de Ada 1roxi duarteNo ratings yet
- Jornada de Sanación y Bendición de Utero PDFDocument4 pagesJornada de Sanación y Bendición de Utero PDFGiselda100% (1)
- Clases 5 - 6 Biologia - CUI UNIVERSIDAD NACIONAL DE TUCUMANDocument7 pagesClases 5 - 6 Biologia - CUI UNIVERSIDAD NACIONAL DE TUCUMANClaudia CampanNo ratings yet
- Gomez Flores Ramos LilianaDocument129 pagesGomez Flores Ramos LilianaAndresPimentelAlvarezNo ratings yet
- 02 Nivel Histologico Protec Mecanicos 2020Document31 pages02 Nivel Histologico Protec Mecanicos 2020estela perezNo ratings yet
- Cuestionario LisosomasDocument4 pagesCuestionario LisosomasgeraldineNo ratings yet
- Cys 25Document69 pagesCys 25AxoncomunicacionNo ratings yet
- Documental sobre bacteriasDocument1 pageDocumental sobre bacteriasPablo Rodriguez100% (3)
- Tema 1Document26 pagesTema 1Dani RondónNo ratings yet