You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Home Remedies EbookDocument357 pagesHome Remedies EbookAbhishek Singh100% (3)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Rife S Fundamental Resonance Enhancement A 100 Effective Cure For CancersDocument28 pagesRife S Fundamental Resonance Enhancement A 100 Effective Cure For CancersDimitar LevskiNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Egg Quality Manual Web PDFDocument44 pagesEgg Quality Manual Web PDFVeterinario La EsperanzaNo ratings yet
- Transpiration POGIL Answer KeyDocument8 pagesTranspiration POGIL Answer KeyJade Tapper50% (2)
- Chiro Guidelines WHODocument51 pagesChiro Guidelines WHOMinh MinhNo ratings yet
- Indian Food Code (25-06-2012)Document60 pagesIndian Food Code (25-06-2012)harry_bccNo ratings yet
- Impact of GST On Unsold Stock of Pre-Packaged CommoditiesDocument2 pagesImpact of GST On Unsold Stock of Pre-Packaged CommoditiesKapil Dev SaggiNo ratings yet
- CDSCO Notice GSTDocument1 pageCDSCO Notice GSTKapil Dev SaggiNo ratings yet
- CC - 60 of 2019Document13 pagesCC - 60 of 2019Kapil Dev SaggiNo ratings yet
- Ceiling Prices of 761 Formulations On Account of GST Implementation.Document49 pagesCeiling Prices of 761 Formulations On Account of GST Implementation.Kapil Dev SaggiNo ratings yet
- LLP Amendment Rules 2016Document4 pagesLLP Amendment Rules 2016Kapil Dev SaggiNo ratings yet
- List of CasesDocument24 pagesList of CasesKapil Dev SaggiNo ratings yet
- Low GST Lawinfotext Blogspot ComDocument3 pagesLow GST Lawinfotext Blogspot ComKapil Dev SaggiNo ratings yet
- NOTICEDocument19 pagesNOTICEPoonkuzhali ManiNo ratings yet
- CBEC Issues Clarification Relating To Waiver of Issuance of SCN and Conclusion of Proceedings in Service Tax and Central ExciseDocument3 pagesCBEC Issues Clarification Relating To Waiver of Issuance of SCN and Conclusion of Proceedings in Service Tax and Central ExciseKapil Dev SaggiNo ratings yet
- Diarmuiid P MeagherDocument333 pagesDiarmuiid P MeagherKapil Dev SaggiNo ratings yet
- GMP in Pharmaceutical IndustryDocument35 pagesGMP in Pharmaceutical IndustrygarcaldecnoNo ratings yet
- Status Report On Start Up IndiaDocument2 pagesStatus Report On Start Up IndiaKapil Dev SaggiNo ratings yet
- Ministry of Railways-Achievements & Initiatives An Ebook by Suresh PrabhuDocument44 pagesMinistry of Railways-Achievements & Initiatives An Ebook by Suresh PrabhuKapil Dev SaggiNo ratings yet
- Modi Govt. AchievementsDocument22 pagesModi Govt. AchievementsKapil Dev SaggiNo ratings yet
- Companies Accounts Second Amendment Rules 2015Document27 pagesCompanies Accounts Second Amendment Rules 2015Kapil Dev SaggiNo ratings yet
- RBI Issues SARFAESI Act 2002 Guidelines and Directions As Amendment Upto 30th June 2015Document26 pagesRBI Issues SARFAESI Act 2002 Guidelines and Directions As Amendment Upto 30th June 2015Kapil Dev SaggiNo ratings yet
- Guidance Note On SEBI Prohibition of Insider Trading Regulations 2015Document3 pagesGuidance Note On SEBI Prohibition of Insider Trading Regulations 2015Kapil Dev SaggiNo ratings yet
- Issue of Shares Under ESOS and Sweat Equity Shares To Persons Resident Outside IndiaDocument8 pagesIssue of Shares Under ESOS and Sweat Equity Shares To Persons Resident Outside IndiaKapil Dev SaggiNo ratings yet
- Format of Regulation 31 of SEBI (Substantial Acquisition of Shares and Takeovers) Regulations, 2011Document3 pagesFormat of Regulation 31 of SEBI (Substantial Acquisition of Shares and Takeovers) Regulations, 2011Shyam SunderNo ratings yet
- CBEC Issues Clarification Relating To Waiver of Issuance of SCN and Conclusion of Proceedings in Service Tax and Central ExciseDocument3 pagesCBEC Issues Clarification Relating To Waiver of Issuance of SCN and Conclusion of Proceedings in Service Tax and Central ExciseKapil Dev SaggiNo ratings yet
- SEBI Specifies Additional Disclosure Requirement by Cos Under Share Based Employee Benefits NormsDocument13 pagesSEBI Specifies Additional Disclosure Requirement by Cos Under Share Based Employee Benefits NormsKapil Dev SaggiNo ratings yet
- FAQs Secretarial AuditDocument19 pagesFAQs Secretarial AuditSaurabh J.No ratings yet
- Master Circular Disclosure in Financial Statements Notes To Accounts'Document40 pagesMaster Circular Disclosure in Financial Statements Notes To Accounts'Kapil Dev SaggiNo ratings yet
- CBDT Notification No. 022015 Dated 13th July 2015Document3 pagesCBDT Notification No. 022015 Dated 13th July 2015Kapil Dev SaggiNo ratings yet
- Bank Finance To Non Banking Financial CompaniesNBFCsDocument13 pagesBank Finance To Non Banking Financial CompaniesNBFCsKapil Dev SaggiNo ratings yet
- Nestle OrderDocument8 pagesNestle OrderNDTVNo ratings yet
- Companies (Amendment) Bill, 2014Document2 pagesCompanies (Amendment) Bill, 2014Kapil Dev SaggiNo ratings yet
- Pemphigus Subtypes Clinical Features Diagnosis andDocument23 pagesPemphigus Subtypes Clinical Features Diagnosis andAnonymous bdFllrgorzNo ratings yet
- Summary of Qualitative Tests (Pharmacognosy)Document8 pagesSummary of Qualitative Tests (Pharmacognosy)kidsaintfineNo ratings yet
- Biology: Pearson EdexcelDocument24 pagesBiology: Pearson EdexcelAhmad MohdNo ratings yet
- Determination of Lethal Dose Ld50of Venom of Four Different Poisonous Snakes Found in PakistanDocument4 pagesDetermination of Lethal Dose Ld50of Venom of Four Different Poisonous Snakes Found in PakistanSutirtho MukherjiNo ratings yet
- 03 Chapter 3Document148 pages03 Chapter 3arghaNo ratings yet
- Cell Membrane and Transport ColoringDocument3 pagesCell Membrane and Transport ColoringTeresa GonzNo ratings yet
- Food Chain QsDocument3 pagesFood Chain QsOliver SansumNo ratings yet
- Cell Division GizmoDocument3 pagesCell Division Gizmoapi-522847737No ratings yet
- Uits PDFDocument36 pagesUits PDFCrystal ParkerNo ratings yet
- Science10 Quarter4 Week3-BiomoleculesDocument2 pagesScience10 Quarter4 Week3-BiomoleculesMargareth LandichoNo ratings yet
- C Specialized GeneralBiology1 Sem1 Clas3 CellsinPlantand-AnimalTissue v2Document14 pagesC Specialized GeneralBiology1 Sem1 Clas3 CellsinPlantand-AnimalTissue v2ダギア メイ アニアNo ratings yet
- General Katalog PT. AmoebaDocument23 pagesGeneral Katalog PT. AmoebaMulyanaNo ratings yet
- Latihan MitosisDocument7 pagesLatihan MitosisChe Mahani HussainNo ratings yet
- Biology ss2Document2 pagesBiology ss2DanielNo ratings yet
- 01 MDCAT SOS Regular Session (5th June-2023) With LR..Document5 pages01 MDCAT SOS Regular Session (5th June-2023) With LR..bakhtawarsrkNo ratings yet
- This Topic. So Congratulations Because Today I'm Going To Talk About TheDocument3 pagesThis Topic. So Congratulations Because Today I'm Going To Talk About TheJoon Bok NamleeNo ratings yet
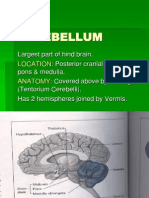
- Physiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamDocument61 pagesPhysiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamZaigham HammadNo ratings yet
- Management of PPHDocument1 pageManagement of PPH098 U.KARTHIK SARAVANA KANTHNo ratings yet
- Human Contact May Reduce Stress in Shelter DogsDocument5 pagesHuman Contact May Reduce Stress in Shelter DogsFlorina AnichitoaeNo ratings yet
- Zoologija VrsteDocument16 pagesZoologija VrsteДушан МарковићNo ratings yet
- Thrombolytic Therapy in Emergency MedicineDocument9 pagesThrombolytic Therapy in Emergency MedicinemedeviNo ratings yet
- MOKUSAKU APPLICATIONS (Version 1)Document1 pageMOKUSAKU APPLICATIONS (Version 1)api-19621568No ratings yet
- Protochordata FIXDocument33 pagesProtochordata FIXSylvia AnggraeniNo ratings yet
- Introduction To Bioinformatics: Database Search (FASTA)Document35 pagesIntroduction To Bioinformatics: Database Search (FASTA)mahedi hasanNo ratings yet
- Ap Environmental Science Free-Response QuestionsDocument6 pagesAp Environmental Science Free-Response Questionsapi-234419767No ratings yet
- Organizing A TextDocument3 pagesOrganizing A TextMercedes Jimenez RomanNo ratings yet