You might also like
- Biochemistry 9th Edition Digital AccessDocument1 pageBiochemistry 9th Edition Digital AccessAlliah CasingNo ratings yet
- Regulación Transcripcional de HTTDocument8 pagesRegulación Transcripcional de HTTHumberto Santana Román100% (1)
- (SUDAH) Kinship Analysis of Offspring of The Giant South American River Turtle (Podocnemis Expansa) Using Microsatellite DNA Markers7Document6 pages(SUDAH) Kinship Analysis of Offspring of The Giant South American River Turtle (Podocnemis Expansa) Using Microsatellite DNA Markers7Takada NamaNo ratings yet
- Nematoide Capilaridae Na Língua de Cerdocyon Thous de Vida Livre No BrasilDocument14 pagesNematoide Capilaridae Na Língua de Cerdocyon Thous de Vida Livre No BrasilMaria EduardaNo ratings yet
- POPULATION GENETICS OF Atta Sexdens Rubropilosa (Hymenoptera: Formicidae)Document16 pagesPOPULATION GENETICS OF Atta Sexdens Rubropilosa (Hymenoptera: Formicidae)Jose Fuentes MarioNo ratings yet
- Research Note / Nota Científica: Haematopinus Tuberculatus Infestation and Distribution inDocument6 pagesResearch Note / Nota Científica: Haematopinus Tuberculatus Infestation and Distribution inNanang AdraverycildNo ratings yet
- Marshall and Berbee 2010 MBEDocument13 pagesMarshall and Berbee 2010 MBEWyth MarshallNo ratings yet
- Paper Herencia ExtracromosomalDocument5 pagesPaper Herencia ExtracromosomalAlexMoretaNo ratings yet
- Characterization of The Estrous Cycle in Galea Spixii (Wagler, 1831)Document6 pagesCharacterization of The Estrous Cycle in Galea Spixii (Wagler, 1831)Haniar MeyNo ratings yet
- Bruno Et Al. (2002) - Nest, Eggs, and Nestlings of The Gray-Headed Tanager (Eucometis Penicillata Albicollis) From São Paulo State, BrazilDocument8 pagesBruno Et Al. (2002) - Nest, Eggs, and Nestlings of The Gray-Headed Tanager (Eucometis Penicillata Albicollis) From São Paulo State, BrazilPaola ResendeNo ratings yet
- Cáceres 2009Document14 pagesCáceres 2009Manu Gustani BussNo ratings yet
- Laboratory Biology of Chetogena Scutellaris (Diptera: Tachinidae), A Parasitoid of Noctuidae, Reared On Fall Armyworm and Cabbage LooperDocument4 pagesLaboratory Biology of Chetogena Scutellaris (Diptera: Tachinidae), A Parasitoid of Noctuidae, Reared On Fall Armyworm and Cabbage Looperromario_ccaNo ratings yet
- 743-Texto Del Artículo-2698-1-10-20200810Document14 pages743-Texto Del Artículo-2698-1-10-20200810Carlos ZaniniNo ratings yet
- Baixa DosDocument9 pagesBaixa DosNathayne SkorupaNo ratings yet
- Ácaros Plumícolas (Acari: Astigmata) em Aves Do Cerrado Do Distrito Federal, BrasilDocument10 pagesÁcaros Plumícolas (Acari: Astigmata) em Aves Do Cerrado Do Distrito Federal, BrasilAnderson FelipeNo ratings yet
- Ectoparasites of Wild Rodents in BrazilDocument7 pagesEctoparasites of Wild Rodents in Braziljuan pablo zuluaga montoyaNo ratings yet
- Telmatobius Helmintos PeruDocument4 pagesTelmatobius Helmintos PeruJorge Manuel Cardenas CallirgosNo ratings yet
- Azpiroz (2015) - Shiny Cowbird (Molothrus Bonariensis) Parasitism Records For Three Globally Threatened Species From The South American PampasDocument8 pagesAzpiroz (2015) - Shiny Cowbird (Molothrus Bonariensis) Parasitism Records For Three Globally Threatened Species From The South American PampasPaola ResendeNo ratings yet
- Litter - Size - and - Embryo - Implantation Silva Et Al 2015 (OA)Document12 pagesLitter - Size - and - Embryo - Implantation Silva Et Al 2015 (OA)Tayssa MarquesNo ratings yet
- Additional Information On Reproductive Behavior of The Red-Breasted Toucan, (Aves: Piciformes: Ramphastidae)Document8 pagesAdditional Information On Reproductive Behavior of The Red-Breasted Toucan, (Aves: Piciformes: Ramphastidae)Miguel ParejoNo ratings yet
- Seasonality, Developing Time and Protandry in Three Populations of The Neotropical Grasshopper Sphenarium Histrio in An Altitudinal GradientDocument21 pagesSeasonality, Developing Time and Protandry in Three Populations of The Neotropical Grasshopper Sphenarium Histrio in An Altitudinal GradientVíctor RamirezdNo ratings yet
- Crawfordl 11 Planirostris in PanamaCity PDFDocument3 pagesCrawfordl 11 Planirostris in PanamaCity PDFAnngkz Franklin AzkabanNo ratings yet
- Additional Novel Cryptosporidium Genotypes in Orname - 2012 - Veterinary ParasitDocument5 pagesAdditional Novel Cryptosporidium Genotypes in Orname - 2012 - Veterinary Parasitfredys seguraNo ratings yet
- 2017 Waichert Et Al More Records PompilidaeDocument7 pages2017 Waichert Et Al More Records PompilidaeZhary VanegasNo ratings yet
- 10 1111@jzs 12337Document28 pages10 1111@jzs 12337Anonymous MXCi0eNo ratings yet
- A Comparison Between Cactophilicy East Communities Isolated From Cereus Hildmannianus and Praecereus Euchlorus Necrotic CladodesDocument9 pagesA Comparison Between Cactophilicy East Communities Isolated From Cereus Hildmannianus and Praecereus Euchlorus Necrotic CladodesBárbara MendonçaNo ratings yet
- García Hernandez Machado 2017 Effectiveness of Maternal Egg Attendance in An Exclusively HarvestmanDocument8 pagesGarcía Hernandez Machado 2017 Effectiveness of Maternal Egg Attendance in An Exclusively Harvestmanlola zavaliaNo ratings yet
- Genetic Determination of The Queen Caste in An Ant Hybrid ZoneDocument4 pagesGenetic Determination of The Queen Caste in An Ant Hybrid ZoneTeresaPeraltaCalvilloNo ratings yet
- Rasoamanana 2019Document12 pagesRasoamanana 2019Daniel Seth AndalNo ratings yet
- Arac 49 02 185Document6 pagesArac 49 02 185sabrina.english.classroomNo ratings yet
- 10 1016@j Actatropica 2006 05 004 PDFDocument3 pages10 1016@j Actatropica 2006 05 004 PDFdianNo ratings yet
- NEW SPECIESDocument6 pagesNEW SPECIESAzhari RizalNo ratings yet
- Kertenkele Canlılığının EvrimiDocument2 pagesKertenkele Canlılığının EvrimiAlara AğalarNo ratings yet
- Morphology, Reproductive Biology and Diet of Colubridae) in Brazilian AmazonDocument7 pagesMorphology, Reproductive Biology and Diet of Colubridae) in Brazilian AmazonEdiegoBatistaNo ratings yet
- Keating EtalDocument9 pagesKeating Etalapi-480736854No ratings yet
- Hernaez Et Al 2010Document8 pagesHernaez Et Al 2010Betel Martinez GuerreroNo ratings yet
- ARC 8 1 (Special Section) 25-32 E79 Low ResDocument8 pagesARC 8 1 (Special Section) 25-32 E79 Low ResAmelia She M UNo ratings yet
- MCR 108Document9 pagesMCR 108Jeahan Farrahdinna A—No ratings yet
- Mapping The Platypus GenomeDocument4 pagesMapping The Platypus GenomeHaynich WilliamsNo ratings yet
- Molecular Genetic Analysis of Male Alternative Strategy and Reproductive Success in The Polygynous Mating BatDocument9 pagesMolecular Genetic Analysis of Male Alternative Strategy and Reproductive Success in The Polygynous Mating BatSiti Rabiatul FajriNo ratings yet
- Barbosa Et Al 2020 - A New Species of Butterfly (Lepidoptera Nymphalidae) From The Highlands of PDFDocument15 pagesBarbosa Et Al 2020 - A New Species of Butterfly (Lepidoptera Nymphalidae) From The Highlands of PDFFernando DiasNo ratings yet
- Effect of Experience on Parasitoid SuccessDocument8 pagesEffect of Experience on Parasitoid SuccessDerrick LeoNo ratings yet
- Urgiles Et Al (2019) Genetic Delimitation of Pristimantis OrestesDocument36 pagesUrgiles Et Al (2019) Genetic Delimitation of Pristimantis OrestesJuan Carlos SánchezNo ratings yet
- Ascaris Lumbricoides Una Revision de Su Epidemiologia y Su Relacion Con Otras InfeccionesDocument12 pagesAscaris Lumbricoides Una Revision de Su Epidemiologia y Su Relacion Con Otras InfeccionesOswaldo GutierrezNo ratings yet
- Quim I Osen SoresDocument9 pagesQuim I Osen Soresnorma patriciaNo ratings yet
- Soneratia AlbaDocument5 pagesSoneratia AlbaRafil FriasmarNo ratings yet
- Von Kossel 2010 (Muscicapidae Luscinia)Document6 pagesVon Kossel 2010 (Muscicapidae Luscinia)Rani Eva DewiNo ratings yet
- Teixeira JR Et Al 2014 A New Two-Pored Amphisbaena Linnaeus, 1758, From Western AmazoniaDocument13 pagesTeixeira JR Et Al 2014 A New Two-Pored Amphisbaena Linnaeus, 1758, From Western AmazoniabasiliscusNo ratings yet
- Fungos No PolénDocument30 pagesFungos No PolénLeticiaNo ratings yet
- A Pluralistic Approach in The Study of Pleurotus Species With Emphasis On Compatibility and Physiology of The European MorphotaxaDocument15 pagesA Pluralistic Approach in The Study of Pleurotus Species With Emphasis On Compatibility and Physiology of The European MorphotaxajakoknoxNo ratings yet
- Gene Expression in The Mixotrophic Prymnesiophyte, Prymnesium, Responds To Prey AvailabilityDocument12 pagesGene Expression in The Mixotrophic Prymnesiophyte, Prymnesium, Responds To Prey AvailabilityAnggie EkaNo ratings yet
- 53 SolanumDocument10 pages53 Solanumvieira.barrosNo ratings yet
- TMP 5 AA9Document8 pagesTMP 5 AA9FrontiersNo ratings yet
- Genetic Diversity of Phil Mallard DuckDocument10 pagesGenetic Diversity of Phil Mallard DuckJudiel DogmaNo ratings yet
- Sexing in Guinea Fowls (Numida Meleagris)Document2 pagesSexing in Guinea Fowls (Numida Meleagris)Nisrina ZhafirahNo ratings yet
- The Queens of The Stingless Bees From Egg To AdultDocument15 pagesThe Queens of The Stingless Bees From Egg To AdultKewen Santiago da Silva LuzNo ratings yet
- baixados (1)Document7 pagesbaixados (1)Nathayne SkorupaNo ratings yet
- Helminth parasites of the Peruvian grunt fish identifiedDocument18 pagesHelminth parasites of the Peruvian grunt fish identifiedDiana PerezNo ratings yet
- PseudanomalaDocument2 pagesPseudanomalacrjesusNo ratings yet
- The Reproductive Ecology of Exotic Trachemys Scripta Elegans in An Invaded Area of Southern EuropeDocument10 pagesThe Reproductive Ecology of Exotic Trachemys Scripta Elegans in An Invaded Area of Southern Europewijayanti chantikaNo ratings yet
- Characterization of Microsatellite Loci of Tetragonisca AngustulaDocument5 pagesCharacterization of Microsatellite Loci of Tetragonisca AngustulaFabiana PiokerNo ratings yet
- Biology, Ecology and Systematics of Australian Scelio: Wasp Parasitoids of Locust and Grasshopper EggsFrom EverandBiology, Ecology and Systematics of Australian Scelio: Wasp Parasitoids of Locust and Grasshopper EggsNo ratings yet
- Ribosomes Structure and FunctionDocument21 pagesRibosomes Structure and FunctionSiddharth BirlaNo ratings yet
- Metagenomics - Remove Host SequencesDocument1 pageMetagenomics - Remove Host SequencesemilioNo ratings yet
- Prenatal Diagnosis FormDocument2 pagesPrenatal Diagnosis FormAnand ChineyNo ratings yet
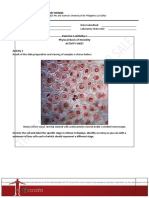
- Exercise 1-Activity 1 Physical Basis of Heredity: Name: Date Submitted: Lab Section: Laboratory InstructorDocument8 pagesExercise 1-Activity 1 Physical Basis of Heredity: Name: Date Submitted: Lab Section: Laboratory InstructorJansen AbadNo ratings yet
- INRA, A New High Frequency Antigen in The INDIAN (IN023) Blood Group SystemDocument4 pagesINRA, A New High Frequency Antigen in The INDIAN (IN023) Blood Group SystemPearlregine Cianne MirandaNo ratings yet
- Complete Genetics Disease ChartDocument14 pagesComplete Genetics Disease ChartJames FlanneryNo ratings yet
- Science Quarter 4-Module 1 Protein Synthesis: NameDocument6 pagesScience Quarter 4-Module 1 Protein Synthesis: NameChristian jade QuijanoNo ratings yet
- (PDF) GenBio2 - Module - Week06Document40 pages(PDF) GenBio2 - Module - Week06Ramon Villota100% (1)
- QP MSC Botany 29012020Document29 pagesQP MSC Botany 29012020kasminpradhaniNo ratings yet
- Training On Molecular Biology, Biotechnology, DNA Fingerprinting and DNA Barcoding Organized by Genomics Research Labs New Delhi India-110092Document9 pagesTraining On Molecular Biology, Biotechnology, DNA Fingerprinting and DNA Barcoding Organized by Genomics Research Labs New Delhi India-110092Escherichia GenomicsNo ratings yet
- Expression Vectors PDFDocument2 pagesExpression Vectors PDFJennifer33% (3)
- Introduction - Phenotypic Plasticity in Development and Evolution - Facts and ConceptsDocument11 pagesIntroduction - Phenotypic Plasticity in Development and Evolution - Facts and ConceptsJose PapoNo ratings yet
- Draft Poster For PSBMBDocument1 pageDraft Poster For PSBMBceejhaybiochemNo ratings yet
- Biology and Evolution of Life ScienceDocument5 pagesBiology and Evolution of Life ScienceAdissa Hasna MutiaraNo ratings yet
- Chapter 9 PDFDocument41 pagesChapter 9 PDFIziz De La CruzNo ratings yet
- Toothpick Fish: A Middle School Activity For Teaching Genetics and Environmental ScienceDocument13 pagesToothpick Fish: A Middle School Activity For Teaching Genetics and Environmental ScienceakgainesNo ratings yet
- B.Sc. BotanyDocument13 pagesB.Sc. Botanyr prathapNo ratings yet
- Science 9Document3 pagesScience 9Hezl Valerie ArzadonNo ratings yet
- Geriatric Physiology by Dr. Leandro D. Vila and Dr. Edmee Y. MartinezDocument72 pagesGeriatric Physiology by Dr. Leandro D. Vila and Dr. Edmee Y. MartinezMelissa SalayogNo ratings yet
- Chlorella SPDocument18 pagesChlorella SPrinifiahNo ratings yet
- Nucleic Acid Hydrolysis TestsDocument5 pagesNucleic Acid Hydrolysis TestsKate de LunaNo ratings yet
- Nonmendelian AssignmentDocument4 pagesNonmendelian AssignmentAnthony HugillNo ratings yet
- Gen. Bio 2 Sex Linkage and RecombinationDocument16 pagesGen. Bio 2 Sex Linkage and RecombinationMeagan TorresNo ratings yet
- The Human Genome ProjectDocument16 pagesThe Human Genome Projectmatt medmedmedicNo ratings yet
- Eukaryotic TranscriptionDocument34 pagesEukaryotic TranscriptionKanaka lata SorenNo ratings yet
- The Significance of MeiosisDocument5 pagesThe Significance of MeiosisKaren CasternoboNo ratings yet
- DNA Fingerprinting: Presented by Pranab Borah Department of Herbal Science & Technology ADP College, NagaonDocument30 pagesDNA Fingerprinting: Presented by Pranab Borah Department of Herbal Science & Technology ADP College, NagaonRavi Poonam Prakash DubeyNo ratings yet
- CH 5Document97 pagesCH 5jaraulabelleNo ratings yet