You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Indigenous EducationDocument475 pagesIndigenous EducationCuentaLibrosScribd100% (1)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Science That Never Has Been CitedDocument4 pagesThe Science That Never Has Been CitedCuentaLibrosScribdNo ratings yet
- High Performance Techniques For Microsoft SQL Server PDFDocument307 pagesHigh Performance Techniques For Microsoft SQL Server PDFmaghnus100% (1)
- Ordinary Dierential Equations Principles and ApplicationsDocument349 pagesOrdinary Dierential Equations Principles and ApplicationsSokratis Spyrou100% (1)
- Blockchain Unconfirmed Transaction Hack Script Docx PDF FreeDocument2 pagesBlockchain Unconfirmed Transaction Hack Script Docx PDF FreeHealing Relaxing Sleep Music100% (1)
- Tonnies Community and Civil SocietyDocument319 pagesTonnies Community and Civil Societydacho146100% (5)
- Elasticity and Its Applications: For Use With Mankiw and Taylor, Economics 4 Edition 9781473725331 © CENGAGE EMEA 2017Document39 pagesElasticity and Its Applications: For Use With Mankiw and Taylor, Economics 4 Edition 9781473725331 © CENGAGE EMEA 2017Joana AgraNo ratings yet
- KUKA Sim 30 Installation enDocument49 pagesKUKA Sim 30 Installation enRégis Naydo0% (1)
- Language Ideologies and Epistemic ExclusionDocument23 pagesLanguage Ideologies and Epistemic ExclusionCuentaLibrosScribdNo ratings yet
- Language Policy and Language IdeologyDocument18 pagesLanguage Policy and Language IdeologyCuentaLibrosScribdNo ratings yet
- Language Ideology and the Prejudice of Standard EnglishDocument16 pagesLanguage Ideology and the Prejudice of Standard EnglishCuentaLibrosScribd100% (1)
- Danaher - Traveller Nomadic and Migrant EducationDocument268 pagesDanaher - Traveller Nomadic and Migrant EducationCuentaLibrosScribdNo ratings yet
- Arc Welding Cracks SolidificationDocument3 pagesArc Welding Cracks SolidificationShaher YarNo ratings yet
- Vendor Information Vishay BLH Handbook TC0013 Solutions For Process Weighing and Force Measurement Electronic Weigh SystemsDocument59 pagesVendor Information Vishay BLH Handbook TC0013 Solutions For Process Weighing and Force Measurement Electronic Weigh SystemsAndrew JacksonNo ratings yet
- Preventing Generator Damage from Hydrogen MoistureDocument27 pagesPreventing Generator Damage from Hydrogen MoistureIskerNo ratings yet
- SVCE Seminar Report Format (FINAL)Document6 pagesSVCE Seminar Report Format (FINAL)Vinod KumarNo ratings yet
- Babcock University Bookshop An Babcock University BookshopDocument11 pagesBabcock University Bookshop An Babcock University BookshopAdeniyi M. AdelekeNo ratings yet
- Atht Model Ques 2017Document1 pageAtht Model Ques 2017Jeyakumar VenugopalNo ratings yet
- IGCSE Chemistry TEST YOUR SELF CHAPTER 4Document12 pagesIGCSE Chemistry TEST YOUR SELF CHAPTER 4Nguyễn Việt Huy RoyNo ratings yet
- Integumentary SystemDocument8 pagesIntegumentary SystemAshley Brithanie RamosNo ratings yet
- XXXXX: Important Instructions To ExaminersDocument21 pagesXXXXX: Important Instructions To ExaminersYogesh DumaneNo ratings yet
- Practical 4 - Signpost 2011Document4 pagesPractical 4 - Signpost 2011Percy Percival50% (2)
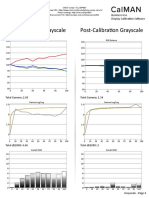
- TCL 55P607 CNET Review Calibration ResultsDocument3 pagesTCL 55P607 CNET Review Calibration ResultsDavid KatzmaierNo ratings yet
- Beyond SVGFDocument66 pagesBeyond SVGFLiliana QueiroloNo ratings yet
- Detailed Geotechnical ReportDocument29 pagesDetailed Geotechnical Reportathar abbasNo ratings yet
- What Is Canal LiningDocument6 pagesWhat Is Canal LiningFiaz GujjarNo ratings yet
- NFRC 200-2010Document45 pagesNFRC 200-2010reynolds534100% (1)
- On Handwritten Digit RecognitionDocument15 pagesOn Handwritten Digit RecognitionAnkit Upadhyay100% (1)
- Historia de CatastroDocument7 pagesHistoria de CatastroMiki Sulca SosaNo ratings yet
- RWM61 Data SheetDocument3 pagesRWM61 Data SheetBarth XaosNo ratings yet
- Refining of MaterialsDocument38 pagesRefining of MaterialsJAWAD AHMAD BURTNo ratings yet
- Nms5Lx 6.6.1: User ManualDocument590 pagesNms5Lx 6.6.1: User ManualGentjan ZogaNo ratings yet
- Dell EMC Avamar NDMP Accelerator For NASDocument58 pagesDell EMC Avamar NDMP Accelerator For NASmanish.puri.gcpNo ratings yet
- Full Text 01Document110 pagesFull Text 01GumbuzaNo ratings yet
- Design of RC Building - ExerciseDocument6 pagesDesign of RC Building - Exercisesajeerala100% (1)
- 2011 Nov P1 Maths L2Document9 pages2011 Nov P1 Maths L2nhlanhlamhlambi3No ratings yet
- Evaluasi Mutu Fisik, Total Bakteri, Dan Sensori Minuman Sari Tempe Dengan Penambahan Bunga KecombrangDocument12 pagesEvaluasi Mutu Fisik, Total Bakteri, Dan Sensori Minuman Sari Tempe Dengan Penambahan Bunga KecombrangJosua PakpahanNo ratings yet