You might also like
- Caso Clinico Cesarea Segmentaria 13-06-16Document39 pagesCaso Clinico Cesarea Segmentaria 13-06-16Wendy Reyes100% (4)
- Plan de Cuidado DiabetesDocument14 pagesPlan de Cuidado DiabetesSandra OlmedaNo ratings yet
- Protocolo Evaluación Clinica Del HABLADocument7 pagesProtocolo Evaluación Clinica Del HABLAMatias OlguinNo ratings yet
- El Milagro de LancianoDocument5 pagesEl Milagro de LancianoTommy ZegarraNo ratings yet
- Metabolismo B Estudiante PrimariaaaaaDocument6 pagesMetabolismo B Estudiante PrimariaaaaaJose Mauro Avila VelaNo ratings yet
- Brunfelsia GrandifloraDocument9 pagesBrunfelsia GrandifloraJose Mauro Avila VelaNo ratings yet
- Biogeo ExpoDocument26 pagesBiogeo ExpoJose Mauro Avila VelaNo ratings yet
- La placenta: órgano vital del embarazoDocument4 pagesLa placenta: órgano vital del embarazoJose Mauro Avila VelaNo ratings yet
- Extracción aceite cedrón métodos vapor hidrodestilaciónDocument30 pagesExtracción aceite cedrón métodos vapor hidrodestilaciónJoel PantojaNo ratings yet
- Vitrificación de Ovocitos Como TécnicaDocument6 pagesVitrificación de Ovocitos Como TécnicaJose Mauro Avila VelaNo ratings yet
- Agroecosistemas Del Valle ChicamaDocument1 pageAgroecosistemas Del Valle ChicamaJose Mauro Avila VelaNo ratings yet
- ORQUIDEASDocument12 pagesORQUIDEASJose Mauro Avila VelaNo ratings yet
- La placenta: órgano vital del embarazoDocument4 pagesLa placenta: órgano vital del embarazoJose Mauro Avila VelaNo ratings yet
- Reproducción en EquinodermosDocument3 pagesReproducción en EquinodermosJose Mauro Avila VelaNo ratings yet
- 2.factores ambientalesCCBB2018Document46 pages2.factores ambientalesCCBB2018Jose Mauro Avila VelaNo ratings yet
- ORQUIDEASDocument12 pagesORQUIDEASJose Mauro Avila VelaNo ratings yet
- Fisio Log I Adela Apoptosis 2008Document56 pagesFisio Log I Adela Apoptosis 2008Ol KeenNo ratings yet
- Cinco Filos de Zoología de InvertebradosDocument9 pagesCinco Filos de Zoología de InvertebradosJose Mauro Avila VelaNo ratings yet
- Sesiones 1 y 2Document36 pagesSesiones 1 y 2Jose Mauro Avila VelaNo ratings yet
- Alumnos Aprobados Curso de Ecología General Vi Ciclo Ciencias Biológicas PDFDocument1 pageAlumnos Aprobados Curso de Ecología General Vi Ciclo Ciencias Biológicas PDFJose Mauro Avila VelaNo ratings yet
- Medidas EstadisticasDocument23 pagesMedidas EstadisticasJose Mauro Avila VelaNo ratings yet
- Medidas EstadisticasDocument23 pagesMedidas EstadisticasJose Mauro Avila VelaNo ratings yet
- Efecto Del PHDocument2 pagesEfecto Del PHJose Mauro Avila VelaNo ratings yet
- Química AnaliticaDocument1 pageQuímica AnaliticaJose Mauro Avila VelaNo ratings yet
- Informe de Biotecnologia 4Document14 pagesInforme de Biotecnologia 4Charles WilsonNo ratings yet
- NitexolDocument27 pagesNitexolAnytta Quilodrán MarquézNo ratings yet
- Prueba 01 Bases Teoricas PsicopedagogiaDocument25 pagesPrueba 01 Bases Teoricas PsicopedagogiaAndrea Martínez Antezana0% (2)
- Producción de OrinaDocument8 pagesProducción de OrinaYadira TrujilloNo ratings yet
- Fichas Miología 1Document15 pagesFichas Miología 1Katherine ScarletNo ratings yet
- ACTIVIDAD 1 MODULO II Desarrollo Durante La AdolescenciaDocument13 pagesACTIVIDAD 1 MODULO II Desarrollo Durante La AdolescenciaYaz ReyezNo ratings yet
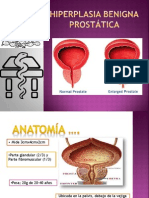
- Hiperplasia Benigna ProstáticaDocument48 pagesHiperplasia Benigna ProstáticaGerardo De La PenaNo ratings yet
- Anestesicos LocalesDocument5 pagesAnestesicos Localesjhony_prieto92No ratings yet
- Productos Fuxion Resumee PDFDocument22 pagesProductos Fuxion Resumee PDFcasacamNo ratings yet
- Síntesis de Ácido MalicoDocument6 pagesSíntesis de Ácido MalicojesusbernalortNo ratings yet
- Renal NormalDocument87 pagesRenal NormalErika MelendezNo ratings yet
- "Intervención Deglutoria en Población Pediátrica": Rodrigo Morales FernándezDocument30 pages"Intervención Deglutoria en Población Pediátrica": Rodrigo Morales FernándezLisbeth Herrera CofréNo ratings yet
- Organos Supra en InframesocolicosDocument11 pagesOrganos Supra en InframesocolicosGeorgina GalindoNo ratings yet
- Medición capacidad vital y flujo aireDocument14 pagesMedición capacidad vital y flujo aireJmontoya RojasNo ratings yet
- Simulación de Paciente AsmáticoDocument6 pagesSimulación de Paciente AsmáticoJecar Vladimir NsNo ratings yet
- Musculo LisoDocument6 pagesMusculo LisoMaurimar LarezNo ratings yet
- Silabos 2015 0 RNR+301Document5 pagesSilabos 2015 0 RNR+301Renzo Terrones MurgaNo ratings yet
- Taller de Bioquimica EnzimasDocument15 pagesTaller de Bioquimica EnzimasAlejandra SalcedoNo ratings yet
- Reporte Cinética Enzimática ACTUAL 2023-1Document9 pagesReporte Cinética Enzimática ACTUAL 2023-1Alta Gracia ArguellesNo ratings yet
- Valores de LaboratorioDocument36 pagesValores de LaboratorioharryparralesNo ratings yet
- Fisiopatologia Del AncianoDocument24 pagesFisiopatologia Del Ancianoluis andres trejo castellanosNo ratings yet
- Absorción y metabolismo del etanol en el cuerpo humanoDocument20 pagesAbsorción y metabolismo del etanol en el cuerpo humanoConnor CocoNo ratings yet
- Garay Rau Alexander Ever Tarea Personal 10Document10 pagesGaray Rau Alexander Ever Tarea Personal 10Thomy Villnueva QuinterosNo ratings yet
- Examen de Biología de la Universidad de Andalucía (2011-2012Document20 pagesExamen de Biología de la Universidad de Andalucía (2011-2012landm1941No ratings yet
- Plan GlobalDocument1 pagePlan Globalapi-543968311No ratings yet
- Cuestionario 2 Bioca CuevasDocument5 pagesCuestionario 2 Bioca CuevasLiriany CuevasNo ratings yet