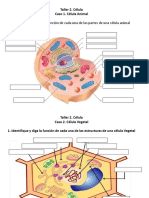
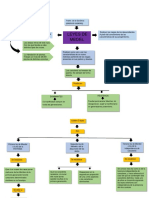
You might also like
- GUIAS GRADO NOVENO MARZO 2020 OkDocument8 pagesGUIAS GRADO NOVENO MARZO 2020 OkYolfranlleNo ratings yet
- Taller Genética Mendeliana.20201Document2 pagesTaller Genética Mendeliana.20201Maria Margarita TisoyNo ratings yet
- Monohíbridos y DihíbridosDocument16 pagesMonohíbridos y DihíbridoskatalinaNo ratings yet
- Guia Didactica Octavo SegundoDocument5 pagesGuia Didactica Octavo SegundoLuis FernandoNo ratings yet
- Las Leyes de MendelDocument3 pagesLas Leyes de MendeldonisemilioNo ratings yet
- La Herencia No Mendeliana para Cuarto de SecundariaDocument3 pagesLa Herencia No Mendeliana para Cuarto de SecundariaAdelmo Jose Herazo Vergara100% (1)
- Práctica de Laboratorio de CromatografíaDocument6 pagesPráctica de Laboratorio de CromatografíaGenesis FloresNo ratings yet
- El NeodarwinismoDocument1 pageEl NeodarwinismoClaudia MarcosNo ratings yet
- Problemas de Genetica2Document4 pagesProblemas de Genetica2JavierNo ratings yet
- Genetica Mendeliana Conjunto de ProblemaDocument55 pagesGenetica Mendeliana Conjunto de ProblemaDanna GonzalezNo ratings yet
- Ejercicios Codominancia y Dominancia IncompletaDocument8 pagesEjercicios Codominancia y Dominancia IncompletaYensy AmadoNo ratings yet
- Tema 2 Taller Parte 1Document6 pagesTema 2 Taller Parte 1alba acuñaNo ratings yet
- Evaluacion Saber Cruces Geneticos 9Document2 pagesEvaluacion Saber Cruces Geneticos 9MARIO CELEDONNo ratings yet
- Guia Iip Biologia Noveno 2021 Darwin Angucho OidorDocument18 pagesGuia Iip Biologia Noveno 2021 Darwin Angucho OidorElColombianoNo ratings yet
- Problemas de Genetica2005Document8 pagesProblemas de Genetica2005Elena Ramonell GarciaNo ratings yet
- Práctica de LaboratorioDocument2 pagesPráctica de LaboratorioRuben FonsecaNo ratings yet
- Boletín InformativoDocument2 pagesBoletín InformativoValentina Michaca GonzalezNo ratings yet
- Clasificacion de Los Seres Vivos Y TaxonomiaDocument19 pagesClasificacion de Los Seres Vivos Y TaxonomiaAndroberto Alva100% (1)
- Problemas de Genética 2 ResueltosDocument6 pagesProblemas de Genética 2 ResueltosYaiza Montero TimonNo ratings yet
- Leyes de Mendel y Patron de Herencia MendelianaDocument4 pagesLeyes de Mendel y Patron de Herencia MendelianaHelena Ferrandis RosellNo ratings yet
- Leyes de MendelDocument2 pagesLeyes de Mendeldaniel100% (1)
- Caracteristica de La PoblacionDocument9 pagesCaracteristica de La PoblacionrosaNo ratings yet
- Guía Laboratorio N°5 EpistasisDocument4 pagesGuía Laboratorio N°5 EpistasisAngelNo ratings yet
- Genetica 2 Ejercicios Cruces1Document1 pageGenetica 2 Ejercicios Cruces1Daniel Sandoval RojasNo ratings yet
- Escuelas TaxonómicasDocument1 pageEscuelas TaxonómicasLiliana RuedaNo ratings yet
- Solucion Problemas GenetcaDocument32 pagesSolucion Problemas Genetcajapylrh100% (1)
- Herencia Del SexoDocument31 pagesHerencia Del SexoPiterVargasHuaman100% (1)
- Examen Final 9 GradoDocument3 pagesExamen Final 9 GradoMeriely SarmientoNo ratings yet
- GUIA Grado Noveno Leyes de MendelDocument4 pagesGUIA Grado Noveno Leyes de MendelCarlos SinitaveNo ratings yet
- Cta Problemas ResueltosDocument12 pagesCta Problemas ResueltosOscar Antonio Moncada VasquezNo ratings yet
- Problem A RioDocument7 pagesProblem A RioKimberly BarretoNo ratings yet
- Taller 1 y 2 de GEN TICA PDFDocument33 pagesTaller 1 y 2 de GEN TICA PDFSolanyi EstupiñanNo ratings yet
- Actividad 5 Genética No MendelianaDocument3 pagesActividad 5 Genética No MendelianaMiguel SilvaNo ratings yet
- Genetica OpcionalDocument18 pagesGenetica OpcionalPaola Gismondi ParedesNo ratings yet
- Problemas de GenéticaDocument8 pagesProblemas de GenéticaAriadna Anthony Vilar100% (1)
- Variabilidad GeneticaDocument45 pagesVariabilidad GeneticaHebert0% (1)
- La Herencia ShowDocument18 pagesLa Herencia Showgrisel_pagánNo ratings yet
- CODOMINANCIADocument2 pagesCODOMINANCIAArturo Gonzalez50% (2)
- Ejercicios GeneticaherenciaDocument11 pagesEjercicios GeneticaherenciaArmando67% (3)
- Problemas Leyes de MendelDocument2 pagesProblemas Leyes de MendelcvenegaslopezNo ratings yet
- La Reproduccion SexualDocument21 pagesLa Reproduccion SexualDamhi LemusNo ratings yet
- Guia 2. Periodo 3. Ecologia de Poblaciones 8Document3 pagesGuia 2. Periodo 3. Ecologia de Poblaciones 8Jose Alberto Leon AldanaNo ratings yet
- Taller GeneticaDocument5 pagesTaller GeneticaJuan David Gomez MeloNo ratings yet
- Relaciones InterespecificasDocument1 pageRelaciones InterespecificasValeria Yepes Moscote100% (2)
- Conceptos Básicos y Leyes de MendelDocument8 pagesConceptos Básicos y Leyes de MendelalegriaaaaNo ratings yet
- Herencia Ligada Al Sexo.Document9 pagesHerencia Ligada Al Sexo.JHONY ANDRES CASTILLA USECHENo ratings yet
- Ejercicios 3 Ley de MendelDocument1 pageEjercicios 3 Ley de MendelDiego AgudeloNo ratings yet
- Problemas Genetica ResueltosDocument15 pagesProblemas Genetica ResueltosNairuxx 128No ratings yet
- 28 Problemas ResueltosDocument15 pages28 Problemas ResueltosClubferrocarril LaDoroteaNo ratings yet
- Desarrollo Embrionario TrabajoDocument5 pagesDesarrollo Embrionario TrabajoVictor Santa Cruz Quiroz100% (2)
- 1a. - Alumno - 2a. Parte - Reproducción y Herencia HumanaDocument4 pages1a. - Alumno - 2a. Parte - Reproducción y Herencia HumanaSandra RodriguezNo ratings yet
- Que Es Una Biomolécula ExposicionDocument4 pagesQue Es Una Biomolécula ExposicionMARIA USTARIZ FRAGOZONo ratings yet
- Ejercicio Leyes de MendelDocument3 pagesEjercicio Leyes de MendelJORGE PORTILLONo ratings yet
- Mapa Genética de PoblacionesDocument1 pageMapa Genética de Poblacioneskaren perez100% (1)
- Desarrollo de Problemas de Genetica MendelianaDocument21 pagesDesarrollo de Problemas de Genetica MendelianaJhazmin Vasquez RodriguezNo ratings yet
- Genética y Transmisión de Caracteres HereditariosDocument23 pagesGenética y Transmisión de Caracteres HereditariosMarelcy calvoNo ratings yet
- Ejercicios de Herencia IntermediaDocument2 pagesEjercicios de Herencia IntermediaSoledad Segovia50% (2)
- Genetica EjerciciosDocument16 pagesGenetica EjerciciosJim Quinde Moreno100% (2)
- Taller de Recuperacion de Grado 9Document6 pagesTaller de Recuperacion de Grado 9maria correa zuñigaNo ratings yet
- Genética y HerenciaDocument4 pagesGenética y Herenciatinamtz082520No ratings yet
- Guia 3 Fisica GeneralDocument4 pagesGuia 3 Fisica GeneralATC26No ratings yet
- Biologia Comun - Aparato DigestivoDocument20 pagesBiologia Comun - Aparato DigestivoKaedre100% (1)
- 07 AdiccionesDocument20 pages07 AdiccionesYonatan DonosoNo ratings yet
- Lactancia MaternaDocument240 pagesLactancia Maternaministeriodesalud100% (1)
- 06 Conducta Suicida PDFDocument7 pages06 Conducta Suicida PDFKaedreNo ratings yet
- Causalidad PsiquiatriaDocument9 pagesCausalidad PsiquiatriaKaedreNo ratings yet
- Investigacion FinalDocument1 pageInvestigacion FinalKaedreNo ratings yet
- Informe 3 Narrativa Detectivesca PDFDocument10 pagesInforme 3 Narrativa Detectivesca PDFKaedreNo ratings yet
- Grado Medicina 2-5Document8 pagesGrado Medicina 2-5KaedreNo ratings yet
- Consejeria MedicaDocument7 pagesConsejeria MedicaKaedreNo ratings yet
- Guia para Progresar Como CorredorDocument382 pagesGuia para Progresar Como CorredorKaedreNo ratings yet
- Biologia DROGASDocument18 pagesBiologia DROGASKaedreNo ratings yet
- Tarea 2, Psicologia Del DesarrolloDocument8 pagesTarea 2, Psicologia Del DesarrolloOrelvi SantosNo ratings yet
- Lab AjoDocument5 pagesLab Ajolaura QuinteroNo ratings yet
- Tema 16. Fisiología Del Sistema Endocrino. IntroducciónDocument31 pagesTema 16. Fisiología Del Sistema Endocrino. IntroducciónRodrigo Eugenio Pérez PortilloNo ratings yet
- Mapa Conceptual de Cambios Reversibles e Irreversible, Tipos de Necrosis y Acumulaciones.Document4 pagesMapa Conceptual de Cambios Reversibles e Irreversible, Tipos de Necrosis y Acumulaciones.Joyce GomezNo ratings yet
- Replicacion de ADNDocument37 pagesReplicacion de ADNNela Palomino LinaresNo ratings yet
- INMUNIZACIONESDocument23 pagesINMUNIZACIONESAlejandroNo ratings yet
- Teoría Endosimbiótica y Adquisición de Organelos CelularesDocument3 pagesTeoría Endosimbiótica y Adquisición de Organelos CelularesMarelcy calvoNo ratings yet
- Acido NucleicoDocument13 pagesAcido NucleicoPhaola GarciaNo ratings yet
- Acidos NucleicosDocument45 pagesAcidos NucleicosJORDY50% (2)
- Biologia FinalizadoDocument11 pagesBiologia FinalizadoMina ARMYNo ratings yet
- Exitus ProtocoloDocument4 pagesExitus ProtocolotirmasultanNo ratings yet
- Taller 9 Las VacunasDocument3 pagesTaller 9 Las VacunasbiologiacolrosarioNo ratings yet
- Clase EpigeneticaDocument61 pagesClase EpigeneticaAmyra Ali AzamarNo ratings yet
- Acidos NucleicosDocument17 pagesAcidos NucleicosMariangelly ChacónNo ratings yet
- Actividad 2. Es Posible La Clonación TerapéuticaDocument3 pagesActividad 2. Es Posible La Clonación Terapéuticapedromendezalvaradoholamsn.com33% (3)
- Tema 6. Ácidos NucleicosDocument12 pagesTema 6. Ácidos NucleicosSantiNo ratings yet
- 1.1 - Caracteristicas Generales de Los ParasitosDocument5 pages1.1 - Caracteristicas Generales de Los ParasitososvaldodosNo ratings yet
- Anatomia y Fisiologia BasicaDocument4 pagesAnatomia y Fisiologia BasicaamethystbaodaiNo ratings yet
- 1.4 HomeostasisDocument10 pages1.4 HomeostasisMayela StnnNo ratings yet
- Bioquímica - RTVDocument19 pagesBioquímica - RTVDaniel GOjedaNo ratings yet
- INFORME DE GENETICA Sobre Desnaturaizacion e hibridacionNNNDocument34 pagesINFORME DE GENETICA Sobre Desnaturaizacion e hibridacionNNNRufino Ttito ApazaNo ratings yet
- Anuario Rec Act Salud 2014Document165 pagesAnuario Rec Act Salud 2014Alan BlondNo ratings yet
- Ficha 4º Semana 5Document4 pagesFicha 4º Semana 5Geraldine Taipe FloresNo ratings yet
- Bloque 1: Temas 1-6Document44 pagesBloque 1: Temas 1-6EstherNo ratings yet
- Lesion CelularDocument3 pagesLesion CelularDaniel CcolccaNo ratings yet
- Estructura, Funcion y Propiedades Del Material GeneticoDocument11 pagesEstructura, Funcion y Propiedades Del Material GeneticoAnonymous pRgQk1Dyi50% (8)
- La Teoría Cromosómica de La HerenciaDocument3 pagesLa Teoría Cromosómica de La HerenciaReineiro GarciaNo ratings yet
- Ciclo de La Tarea 5 - 152001 - 1.Document13 pagesCiclo de La Tarea 5 - 152001 - 1.yuli paolaNo ratings yet
- Medicina Forense Aplicaciones Teorico-Practicas GrandiniDocument44 pagesMedicina Forense Aplicaciones Teorico-Practicas GrandinidavidNo ratings yet
- Sindrome de PearsonDocument8 pagesSindrome de PearsonJesús Sánchez G.No ratings yet